Effect of potential pathogenic gene PDX1 variants of total anomalous pulmonary venous connection on its gene function
FENG Weiqi,, ZHANG Qi, WU Yizhuo, LU Yanan, YU Yu,
Institute for Developmental and Regenerative Cardiovascular Medicine, Xinhua Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai 200092, China
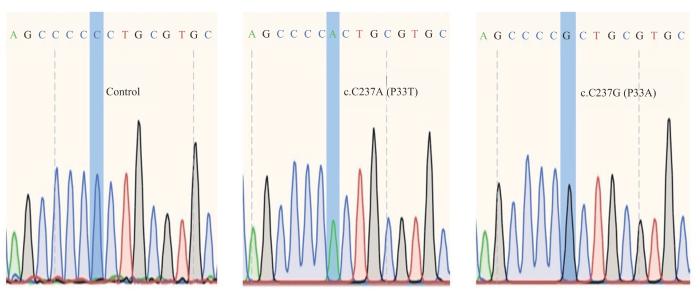
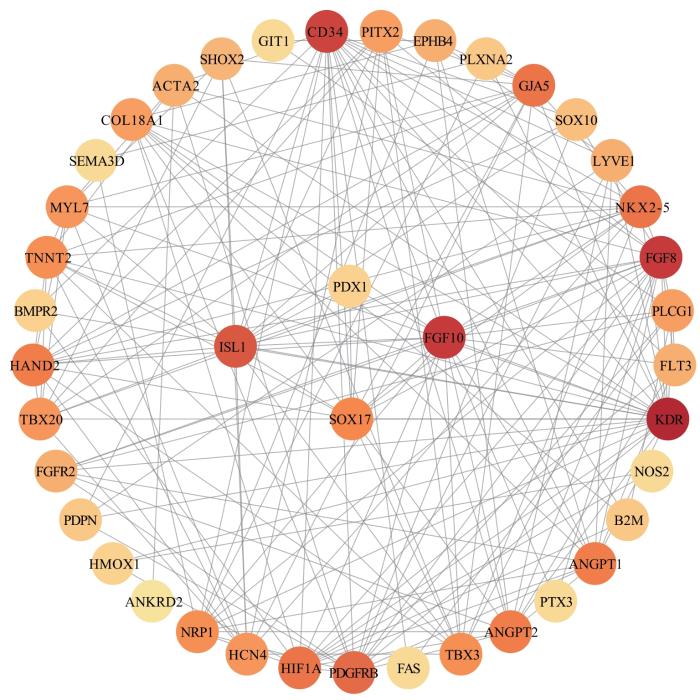
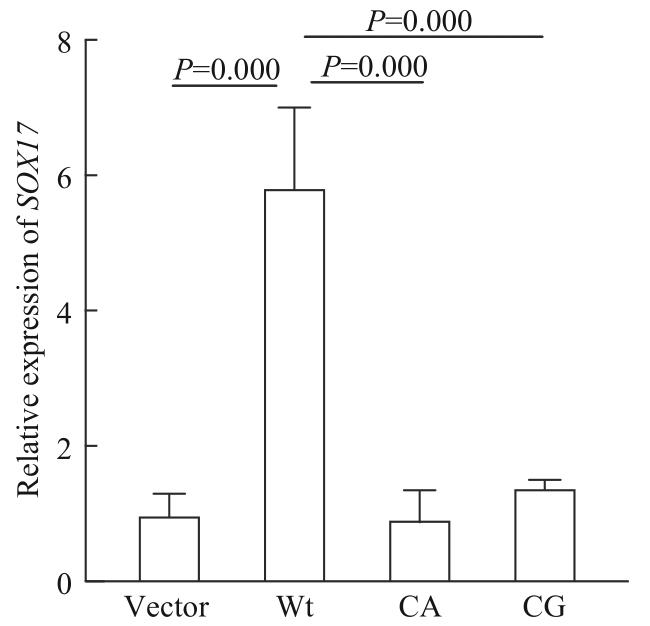
Objective ·To explore the possible pathogenic gene of total anomalous pulmonary venous connection (TAPVC) by whole exon sequencing and verify its function. Methods ·One hundred TAPVC children (case group) and one hundred and twenty healthy children (control group) in Xinhua Hospital and Shanghai Children's Medical Center, Shanghai Jiao Tong University School of Medicine from 2014 to 2019 were included. The blood samples from the two groups of children were collected, and whole blood genomic DNA was extracted for exon sequencing to screen out the potential pathogenic genes of TAPVC. Harmful mutation sites of pathogenic genes were screened through Mutation Taster, SIFT and PolyPhen-2 websites, and then conducted by Sanger sequencing. The wild-type (wild-type group) and mutant (mutant group) plasmids of PDX1 were transfected into HUVEC cells. Quantitative real-time PCR (qPCR) and Western blotting were used to detect the effects of mutations on mRNA and protein levels of PDX1, respectively. The STRING database was used to analyze the interaction between proteins, and qPCR was used to determine the expressions of downstream genes regulated by PDX1. Results ·Pathogenic PDX1 was found in TAPVC children, and Sanger sequencing revealed two novel variants in the gene: c.C237A (P33T) and c. C237G (P33A). Compared with the wild-type group, there was no significant difference in PDX1 mRNA levels in the two mutant groups, but there was a significant increase in relative protein expression of the CA group and CG group, which was 2.9 and 3.4 times higher than the wild-type group, respectively (P=0.000, P=0.001). Protein interaction analysis demonstrated that PDX1 was associated with SOX17. qPCR results showed that overexpression of PDX1 could downregulate the expression of SOX17 in HUVEC. Conclusion ·The two novel PDX1 missense mutations can affect the process of PDX1 post-transcriptional translation, indicating that PDX1 may participate in the occurrence and development of TAPVC by regulating SOX17.
FENG Weiqi, ZHANG Qi, WU Yizhuo, LU Yanan, YU Yu. Effect of potential pathogenic gene PDX1 variants of total anomalous pulmonary venous connection on its gene function. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(10): 1219-1226 doi:10.3969/j.issn.1674-8115.2023.10.001
Note:A. Detection of relative expression of PDX1 in HUVECs by qPCR. B. Detection of the expression of PDX1 in HUVECs by Western blotting. C. Quantitative analysis of the expression of PDX1.
Fig 3
Effect of two mutations of PDX1 on its mRNA and protein expression
在本研究中,我们分别收集了78例TAPVC患儿和100例健康儿童的血液样本,抽提基因组DNA并行WES。通过对WES数据进行SNP关联分析、Fisher精确检验及基因负荷检验,我们筛选出了TAPVC可能的致病基因——PDX1,这是本课题组前期研究[23]已报道的致病基因ARHGEF16以外的、新发现的可能与TAPVC相关的候选基因。作为胰腺β细胞分化和成熟所必需的关键调节因子,PDX1是调节β细胞中的胰岛素基因表达的关键因子[24],且PDX1的杂合突变与糖尿病的发生有关,包括Ⅳ型成熟期糖尿病(maturity-onset diabetes of the young Ⅳ,MODY Ⅳ)和非MODY Ⅱ型糖尿病。
The study was designed by FENG Weiqi, ZHANG Qi and YU Yu. The manuscript was drafted and revised by FENG Weiqi, WU Yizhuo, LU Yanan and YU Yu. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
All authors disclose no relevant conflict of interests.
KAO C C, HSIEH C C, CHENG P J, et al. Total anomalous pulmonary venous connection: from embryology to a prenatal ultrasound diagnostic update[J]. J Med Ultrasound, 2017, 25(3): 130-137.
PALADINI D, PISTORIO A, WU L H, et al. Prenatal diagnosis of total and partial anomalous pulmonary venous connection: multicenter cohort study and meta-analysis[J]. Ultrasound Obstet Gynecol, 2018, 52(1): 24-34.
RELLER M D, STRICKLAND M J, RIEHLE-COLARUSSO T, et al. Prevalence of congenital heart defects in metropolitan Atlanta, 1998‒2005[J]. J Pediatr, 2008, 153(6): 807-813.
CRAIG J M, DARLING R C, ROTHNEY W B. Total pulmonary venous drainage into the right side of the heart; report of 17 autopsied cases not associated with other major cardiovascular anomalies[J]. Lab Invest, 1957, 6(1): 44-64.
NEILL C A, FERENCZ C, SABISTON D C, et al. The familial occurrence of hypoplastic right lung with systemic arterial supply and venous drainage "scimitar syndrome"[J]. Bull Johns Hopkins Hosp, 1960, 107: 1-21.
LI J, YANG S W, PU Z N, et al. Whole-exome sequencing identifies SGCD and ACVRL1 mutations associated with total anomalous pulmonary venous return (TAPVR) in Chinese population[J]. Oncotarget, 2017, 8(17): 27812-27819.
CINQUETTI R, BADI I, CAMPIONE M, et al. Transcriptional deregulation and a missense mutation define ANKRD1 as a candidate gene for total anomalous pulmonary venous return[J]. Hum Mutat, 2008, 29(4): 468-474.
HOLLAND A M, HALE M A, KAGAMI H, et al. Experimental control of pancreatic development and maintenance[J]. Proc Natl Acad Sci U S A, 2002, 99(19): 12236-12241.
STOFFERS D A, ZINKIN N T, STANOJEVIC V, et al. Pancreatic agenesis attributable to a single nucleotide deletion in the human IPF1 gene coding sequence[J]. Nat Genet, 1997, 15(1): 106-110.
GÜRSON C T, TAHSINOGLU M, YAKACIKLI S, et al. A case of agenesis of the dorsal pancreas with interventricular septal defect in an infant[J]. Helv Paediatr Acta, 1970, 25(5): 522-526.
YORIFUJI T, MATSUMURA M, OKUNO T, et al. Hereditary pancreatic hypoplasia, diabetes mellitus, and congenital heart disease: a new syndrome?[J]. J Med Genet, 1994, 31(4): 331-333.
D'AMATO E, GIACOPELLI F, GIANNATTASIO A, et al. Genetic investigation in an Italian child with an unusual association of atrial septal defect, attributable to a new familial GATA4 gene mutation, and neonatal diabetes due to pancreatic agenesis[J]. Diabet Med, 2010, 27(10): 1195-1200.
BHUSHAN A, ITOH N, KATO S, et al. Fgf10 is essential for maintaining the proliferative capacity of epithelial progenitor cells during early pancreatic organogenesis[J]. Development, 2001, 128(24): 5109-5117.
WANG W P, SHI Q, GUO T, et al. PDX1 and ISL1 differentially coordinate with epigenetic modifications to regulate insulin gene expression in varied glucose concentrations[J]. Mol Cell Endocrinol, 2016, 428: 38-48.
WU Y, WHARTON J, WALTERS R, et al. The pathophysiological role of novel pulmonary arterial hypertension gene SOX17[J]. Eur Respir J, 2021, 58(3): 2004172.
HE X, SEMENOV M, TAMAI K, et al. LDL receptor-related proteins 5 and 6 in Wnt/β-catenin signaling: arrows point the way[J]. Development, 2004, 131(8): 1663-1677.
DEGENHARDT K, SINGH M K, AGHAJANIAN H, et al. Semaphorin 3d signaling defects are associated with anomalous pulmonary venous connections[J]. Nat Med, 2013, 19(6): 760-765.
WANG J, JIN L H, ZHANG Q, et al. ARHGEF16 variants screening and mutation function analysis for children with total anomalous pulmonary venous connection[J]. J Shanghai Jiao Tong Univ (Med Sci), 2020, 40(1): 70-75
... 在本研究中,我们分别收集了78例TAPVC患儿和100例健康儿童的血液样本,抽提基因组DNA并行WES.通过对WES数据进行SNP关联分析、Fisher精确检验及基因负荷检验,我们筛选出了TAPVC可能的致病基因——PDX1,这是本课题组前期研究[23]已报道的致病基因ARHGEF16以外的、新发现的可能与TAPVC相关的候选基因.作为胰腺β细胞分化和成熟所必需的关键调节因子,PDX1是调节β细胞中的胰岛素基因表达的关键因子[24],且PDX1的杂合突变与糖尿病的发生有关,包括Ⅳ型成熟期糖尿病(maturity-onset diabetes of the young Ⅳ,MODY Ⅳ)和非MODY Ⅱ型糖尿病. ...
1
... 在本研究中,我们分别收集了78例TAPVC患儿和100例健康儿童的血液样本,抽提基因组DNA并行WES.通过对WES数据进行SNP关联分析、Fisher精确检验及基因负荷检验,我们筛选出了TAPVC可能的致病基因——PDX1,这是本课题组前期研究[23]已报道的致病基因ARHGEF16以外的、新发现的可能与TAPVC相关的候选基因.作为胰腺β细胞分化和成熟所必需的关键调节因子,PDX1是调节β细胞中的胰岛素基因表达的关键因子[24],且PDX1的杂合突变与糖尿病的发生有关,包括Ⅳ型成熟期糖尿病(maturity-onset diabetes of the young Ⅳ,MODY Ⅳ)和非MODY Ⅱ型糖尿病. ...
1
... 在本研究中,我们分别收集了78例TAPVC患儿和100例健康儿童的血液样本,抽提基因组DNA并行WES.通过对WES数据进行SNP关联分析、Fisher精确检验及基因负荷检验,我们筛选出了TAPVC可能的致病基因——PDX1,这是本课题组前期研究[23]已报道的致病基因ARHGEF16以外的、新发现的可能与TAPVC相关的候选基因.作为胰腺β细胞分化和成熟所必需的关键调节因子,PDX1是调节β细胞中的胰岛素基因表达的关键因子[24],且PDX1的杂合突变与糖尿病的发生有关,包括Ⅳ型成熟期糖尿病(maturity-onset diabetes of the young Ⅳ,MODY Ⅳ)和非MODY Ⅱ型糖尿病. ...





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}