Pyroptosis, a new mode of programmed cell death, is primarily characterized by persistent cellular swelling that culminates in cell rupture. This process results in the release of large amounts of inflammatory factors, subsequently triggering an inflammatory response. Benign prostatic hyperplasia (BPH) is the most frequent urological disease in old males and is closely associated with changes in hormones and inflammation response. In recent years, the role of pyroptosis in the occurrence and development of BPH has also received increasing attention. This article summarizes the mechanisms of pyroptosis, concludes the pathogenesis associated with BPH in old males, and outlines the role of pyroptosis in BPH, to provide new ideas for finding more effective therapeutic measures for BPH through pyroptosis.
CHEN Zixuan, LI Dong, LIU Min. Review of the role of pyroptosis in benign prostatic hyperplasia in old males. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(12): 1569-1576 doi:10.3969/j.issn.1674-8115.2023.12.013
细胞焦亡是一种由炎性小体激活、Gasdermin蛋白介导的溶解性和炎症性细胞程序性死亡[7]。具体来说,炎症小体能激活含半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,caspase)家族的部分蛋白(如caspase-1/3/4/5/11),激活后的caspase蛋白通过切割Gasdermin蛋白使之活化。随后,活化的Gasdermin蛋白转位到质膜并在膜上形成孔隙,导致膜破裂、细胞肿胀溶解、炎症因子和微泡等细胞内容物释放,最终引发炎症反应[8-9]。在上述过程中,根据被激活的caspase蛋白的不同,引发的细胞焦亡途径也不尽相同。目前,细胞焦亡的发生主要为依赖caspase-1、非依赖caspase-1共2种途径。
1.1 依赖caspase-1途径
细胞焦亡的依赖caspase-1途径又称经典途径。在病原体相关分子模式和损伤相关分子模式(pathogen-associated molecular patterns and damage-associated molecular patterns,PAMPs and DAMPs)的刺激下,胞浆内的细胞质模式识别受体(pattern recognition receptors,PRRs)被激活[10]。在激活的PRRs的刺激下,caspase-1前体可被含有caspase募集结构域(caspase recruitment domain,CARD)的PRRs直接募集或经含CARD结构的凋亡相关斑点样蛋白(apoptosis-associated speck-like protein containing a CARD,ASC)间接募集,共同组装为caspase-1依赖性炎性小体。随后,该小体可通过自裂激活为caspase-1。而后,caspase-1以如下2种方式引起细胞焦亡:①将Gasdermin蛋白D(Gasdermin-D,GSDMD)裂解为N-GSDMD。后者能穿透细胞膜并在膜上形成孔隙,引起细胞渗透压改变,从而导致细胞肿胀至破裂。②把白细胞介素-1β(interleukin-1β,IL-1β)、IL-18的前体裂解为成熟的IL-1β、IL-18,引发炎症反应[11]。
在上述过程中,PRRs是细胞焦亡启动的关键。作为PRRs的主要组成部分,NOD样受体(NOD-like receptors,NLRs)也在细胞焦亡的启动中扮演了重要角色。常见的NLRs有NOD样受体热蛋白结构域相关蛋白1(NOD-like receptor thermal protein domain associated protein 1,NLRP1)、NLRP3、含caspase激活和募集结构域的NOD样受体4(NOD-like receptor containing a caspase activating and recruitment domain 4,NLRC4)、NLRP6和NLRP9b,它们均以不同的方式参与了细胞焦亡的启动[12]:①NLRP1通过特异性识别炭疽芽孢杆菌的致死因子蛋白酶、福氏志贺菌分泌的泛素连接酶等,激活caspase-1后启动细胞焦亡[13]。②NLRP3在受到微生物、颗粒物质(如错误折叠的蛋白质)和损伤相关分子(如细胞外ATP)等的激活后,可诱导细胞焦亡的发生[14],如细胞外ATP通过激活P2X嘌呤受体7(P2X7)引起K+外排,进而激活NLRP3并促使IL-1β分泌及caspase-1活化,启动细胞焦亡的依赖caspase-1途径[11]。③NLRC4炎性小体的NLR家族凋亡抑制蛋白(NLR family apoptosis inhibitory protein,NAIP)结构在特异性识别PAMPs后,可形成NLRC4炎性小体复合物,该复合物通过激活caspase-1来启动细胞焦亡。④NLRP6和NLRP9b主要在肠上皮细胞中表达,与结肠炎的进展相关[15]。前者可通过激活caspase-1来裂解GSDMD,导致细胞焦亡发生;后者既可通过caspase-1裂解GSDMD引发细胞焦亡,又可通过caspase-1将IL-18前体裂解为IL-18来引起炎症反应,最终激活细胞焦亡的经典途径。
The review was designed by CHEN Zixuan and LIU Min. The manuscript was drafted and revised by CHEN Zixuan, LI Dong and LIU Min. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
ZHANG H B, ZHAO Y N, YANG X J. Role and therapeutic implications of pyroptosis in intervertebral disc degeneration[J]. Chinese Journal of Tissue Engineering Research, 2022, 26(9): 1445-1451.
ZHENG X T, CHEN W W, GONG F C, et al. The role and mechanism of pyroptosis and potential therapeutic targets in Sepsis: a review[J]. Front Immunol, 2021, 12: 711939.
NAIYILA X, LI J Z, HUANG Y, et al. A novel insight into the immune-related interaction of inflammatory cytokines in benign prostatic hyperplasia[J]. J Clin Med, 2023, 12(5): 1821.
TRIPATHI U, MISRA A, TCHKONIA T, et al. Impact of senescent cell subtypes on tissue dysfunction and repair: importance and research questions[J]. Mech Ageing Dev, 2021, 198: 111548.
LI Z A, XIE J J, CHEN J X, et al. Role of microenvironment in the pathogenesis of benign prostatic hyperplasia[J]. Chinese Journal of Urology, 2022, 43(9): 717-720.
ZHAO M, GUO J, GAO Q H, et al. Relationship between pyroptosis-mediated inflammation and the pathogenesis of prostate disease[J]. Front Med (Lausanne), 2023, 10: 1084129.
ZHENG X Y, WANG X Y, ZHANG Y J. Improvement of alveolarization arrest in newborn rats with bronchopulmonary dysplasia via inhibiting alveolar epithelial cell pyroptosis[J]. Journal of Shanghai Jiao tong University:Medical Science, 2023, 43(2): 171-179.
HSU S K, LI C Y, LIN I L, et al. Inflammation-related pyroptosis, a novel programmed cell death pathway, and its crosstalk with immune therapy in cancer treatment[J]. Theranostics, 2021, 11(18): 8813-8835.
MA Q. Pharmacological inhibition of the NLRP3 inflammasome: structure, molecular activation, and inhibitor-NLRP3 interaction[J]. Pharmacol Rev, 2023, 75(3): 487-520.
ZHOU Z W, HE H B, WANG K, et al. Granzyme A from cytotoxic lymphocytes cleaves GSDMB to trigger pyroptosis in target cells[J]. Science, 2020, 368(6494): eaaz7548.
SARRIÓ D, MARTÍNEZ-VAL J, MOLINA-CRESPO Á, et al. The multifaceted roles of gasdermins in cancer biology and oncologic therapies[J]. Biochim Biophys Acta Rev Cancer, 2021, 1876(2): 188635.
OLTRA S S, COLOMO S, SIN L, et al. Distinct GSDMB protein isoforms and protease cleavage processes differentially control pyroptotic cell death and mitochondrial damage in cancer cells[J]. Cell Death Differ, 2023, 30(5): 1366-1381.
JIN B R, KIM H J, NA J H, et al. Targeting benign prostate hyperplasia treatments: AR/TGF-β/NOX4 inhibition by apocynin suppresses inflammation and proliferation[J]. J Adv Res, 2023: S2090-S1232(23)00112-1.
TONG Y, ZHOU R Y. Review of the roles and interaction of androgen and inflammation in benign prostatic hyperplasia[J]. Mediators Inflamm, 2020, 2020: 7958316.
HONG G L, KIM K H, KIM Y J, et al. Decreased mitophagy aggravates benign prostatic hyperplasia in aged mice through DRP1 and estrogen receptor α[J]. Life Sci, 2022, 309: 120980.
CANNARELLA R, CONDORELLI R A, BARBAGALLO F, et al. Endocrinology of the aging prostate: current concepts[J]. Front Endocrinol (Lausanne), 2021, 12: 554078.
AHEARN T U, PEISCH S, PETTERSSON A, et al. Expression of IGF/insulin receptor in prostate cancer tissue and progression to lethal disease[J]. Carcinogenesis, 2018, 39(12): 1431-1437.
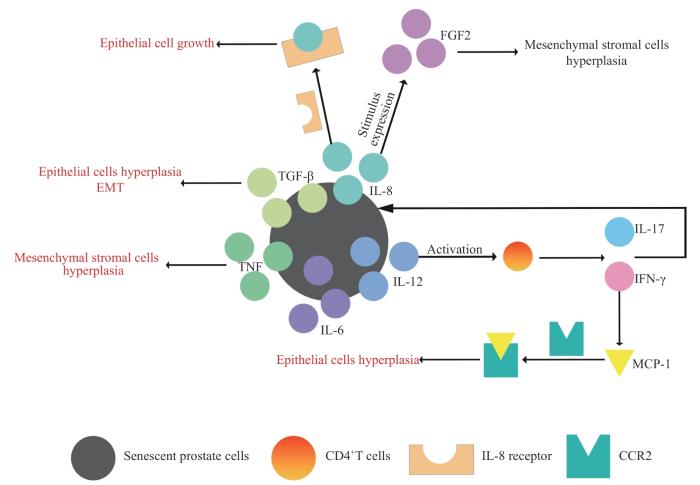
CAO D H, SUN R N, PENG L, et al. Immune cell proinflammatory microenvironment and androgen-related metabolic regulation during benign prostatic hyperplasia in aging[J]. Front Immunol, 2022, 13: 842008.
FIARD G, STAVRINIDES V, CHAMBERS E S, et al. Cellular senescence as a possible link between prostate diseases of the ageing male[J]. Nat Rev Urol, 2021, 18(10): 597-610.
CAO Y, ZHANG H, TU G L, et al. The symptoms of benign prostatic hyperplasia patients with stromal-dominated hyperplasia nodules may be associated with prostate fibrosis[J]. Int J Gen Med, 2023, 16: 1181-1191.
ROYUELA M, DE MIGUEL M P, BETHENCOURT F R, et al. Transforming growth factor beta 1 and its receptor types Ⅰ and Ⅱ. Comparison in human normal prostate, benign prostatic hyperplasia, and prostatic carcinoma[J]. Growth Factors, 1998, 16(2): 101-110.
VICKMAN R E, AARON-BROOKS L, ZHANG R Y, et al. TNF is a potential therapeutic target to suppress prostatic inflammation and hyperplasia in autoimmune disease[J]. Nat Commun, 2022, 13(1): 2133.
ALONSO-MAGDALENA P, BRÖSSNER C, REINER A, et al. A role for epithelial-mesenchymal transition in the etiology of benign prostatic hyperplasia[J]. Proc Natl Acad Sci USA, 2009, 106(8): 2859-2863.
LI Q, HONG Y F, CHEN J, et al. Hypoxia-induced HIF-1α expression promotes neurogenic bladder fibrosis via EMT and pyroptosis[J]. Cells, 2022, 11(23): 3836.
WANG Z, ZHANG Y C, ZHAO C, et al. The miR-223-3p/MAP1B axis aggravates TGF-β-induced proliferation and migration of BPH-1 cells[J]. Cell Signal, 2021, 84: 110004.
JIA C Q, ZHANG Z Q, TANG J, et al. Epithelial-mesenchymal transition induces GSDME transcriptional activation for inflammatory pyroptosis[J]. Front Cell Dev Biol, 2021, 9: 781365.
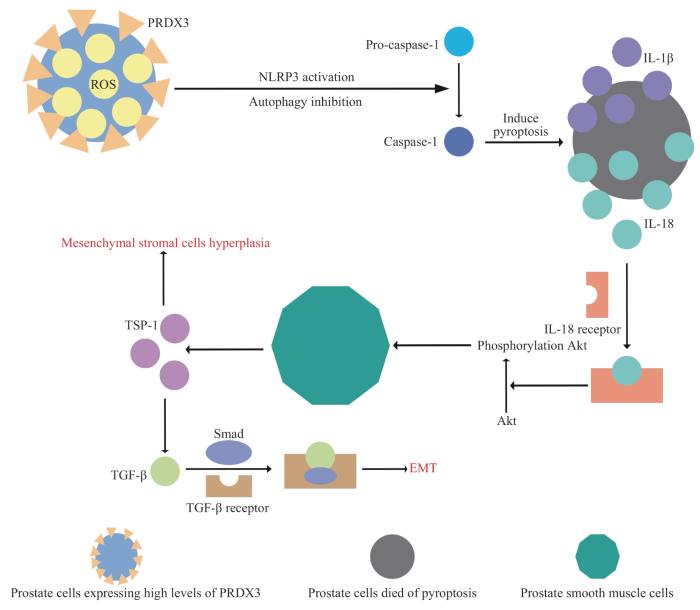
ZHANG C, ZHAI T Y, ZHU J H, et al. Research progress of antioxidants in oxidative stress therapy after spinal cord injury[J]. Neurochem Res, 2023, 48(12): 3473-3484.
MIAO C Y, ZHAO Y, CHEN Y, et al. Investigation of He's Yang Chao recipe against oxidative stress-related mitophagy and pyroptosis to improve ovarian function[J]. Front Endocrinol (Lausanne), 2023, 14: 1077315.
ZHANG C Y, LIN T J, NIE G H, et al. Cadmium and molybdenum co-induce pyroptosis via ROS/PTEN/PI3K/AKT axis in duck renal tubular epithelial cells[J]. Environ Pollut, 2021, 272: 116403.
JIANG M Y, HAN Z D, LI W J, et al. Mitochondrion-associated protein peroxiredoxin 3 promotes benign prostatic hyperplasia through autophagy suppression and pyroptosis activation[J]. Oncotarget, 2017, 8(46): 80295-80302.
QUAN Y, XIN Y G, TIAN G E, et al. Mitochondrial ROS-modulated mtDNA: a potential target for cardiac aging[J]. Oxid Med Cell Longev, 2020, 2020: 9423593.
CHEN W F, HUANG X Q, PENG A X, et al. Kangquan recipe regulates the expression of BAMBI protein via the TGF- β/Smad signaling pathway to inhibit benign prostatic hyperplasia in rats[J]. Evid Based Complement Alternat Med, 2019, 2019: 6281819.
FUSCO F, CRETA M, DE NUNZIO C, et al. Progressive bladder remodeling due to bladder outlet obstruction: a systematic review of morphological and molecular evidences in humans[J]. BMC Urol, 2018, 18(1): 15.
WANG K, CHEN L, YANG J, et al. Urethral meatus stricture BOO stimulates bladder smooth muscle cell proliferation and pyroptosis via IL‑1β and the SGK1‑NFAT2 signaling pathway[J]. Mol Med Rep, 2020, 22(1): 219-226.
KUSTRIMOVIC N, BOMBELLI R, BACI D, et al. Microbiome and prostate cancer: a novel target for prevention and treatment[J]. Int J Mol Sci, 2023, 24(2): 1511.
BERTHELOOT D, LATZ E, FRANKLIN B S. Necroptosis, pyroptosis and apoptosis: an intricate game of cell death[J]. Cell Mol Immunol, 2021, 18(5): 1106-1121.
... 细胞焦亡是一种由炎性小体激活、Gasdermin蛋白介导的溶解性和炎症性细胞程序性死亡[7].具体来说,炎症小体能激活含半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,caspase)家族的部分蛋白(如caspase-1/3/4/5/11),激活后的caspase蛋白通过切割Gasdermin蛋白使之活化.随后,活化的Gasdermin蛋白转位到质膜并在膜上形成孔隙,导致膜破裂、细胞肿胀溶解、炎症因子和微泡等细胞内容物释放,最终引发炎症反应[8-9].在上述过程中,根据被激活的caspase蛋白的不同,引发的细胞焦亡途径也不尽相同.目前,细胞焦亡的发生主要为依赖caspase-1、非依赖caspase-1共2种途径. ...
1
... 细胞焦亡是一种由炎性小体激活、Gasdermin蛋白介导的溶解性和炎症性细胞程序性死亡[7].具体来说,炎症小体能激活含半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,caspase)家族的部分蛋白(如caspase-1/3/4/5/11),激活后的caspase蛋白通过切割Gasdermin蛋白使之活化.随后,活化的Gasdermin蛋白转位到质膜并在膜上形成孔隙,导致膜破裂、细胞肿胀溶解、炎症因子和微泡等细胞内容物释放,最终引发炎症反应[8-9].在上述过程中,根据被激活的caspase蛋白的不同,引发的细胞焦亡途径也不尽相同.目前,细胞焦亡的发生主要为依赖caspase-1、非依赖caspase-1共2种途径. ...
1
... 细胞焦亡是一种由炎性小体激活、Gasdermin蛋白介导的溶解性和炎症性细胞程序性死亡[7].具体来说,炎症小体能激活含半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,caspase)家族的部分蛋白(如caspase-1/3/4/5/11),激活后的caspase蛋白通过切割Gasdermin蛋白使之活化.随后,活化的Gasdermin蛋白转位到质膜并在膜上形成孔隙,导致膜破裂、细胞肿胀溶解、炎症因子和微泡等细胞内容物释放,最终引发炎症反应[8-9].在上述过程中,根据被激活的caspase蛋白的不同,引发的细胞焦亡途径也不尽相同.目前,细胞焦亡的发生主要为依赖caspase-1、非依赖caspase-1共2种途径. ...
1
... 细胞焦亡是一种由炎性小体激活、Gasdermin蛋白介导的溶解性和炎症性细胞程序性死亡[7].具体来说,炎症小体能激活含半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,caspase)家族的部分蛋白(如caspase-1/3/4/5/11),激活后的caspase蛋白通过切割Gasdermin蛋白使之活化.随后,活化的Gasdermin蛋白转位到质膜并在膜上形成孔隙,导致膜破裂、细胞肿胀溶解、炎症因子和微泡等细胞内容物释放,最终引发炎症反应[8-9].在上述过程中,根据被激活的caspase蛋白的不同,引发的细胞焦亡途径也不尽相同.目前,细胞焦亡的发生主要为依赖caspase-1、非依赖caspase-1共2种途径. ...
1
... 细胞焦亡的依赖caspase-1途径又称经典途径.在病原体相关分子模式和损伤相关分子模式(pathogen-associated molecular patterns and damage-associated molecular patterns,PAMPs and DAMPs)的刺激下,胞浆内的细胞质模式识别受体(pattern recognition receptors,PRRs)被激活[10].在激活的PRRs的刺激下,caspase-1前体可被含有caspase募集结构域(caspase recruitment domain,CARD)的PRRs直接募集或经含CARD结构的凋亡相关斑点样蛋白(apoptosis-associated speck-like protein containing a CARD,ASC)间接募集,共同组装为caspase-1依赖性炎性小体.随后,该小体可通过自裂激活为caspase-1.而后,caspase-1以如下2种方式引起细胞焦亡:①将Gasdermin蛋白D(Gasdermin-D,GSDMD)裂解为N-GSDMD.后者能穿透细胞膜并在膜上形成孔隙,引起细胞渗透压改变,从而导致细胞肿胀至破裂.②把白细胞介素-1β(interleukin-1β,IL-1β)、IL-18的前体裂解为成熟的IL-1β、IL-18,引发炎症反应[11]. ...
2
... 细胞焦亡的依赖caspase-1途径又称经典途径.在病原体相关分子模式和损伤相关分子模式(pathogen-associated molecular patterns and damage-associated molecular patterns,PAMPs and DAMPs)的刺激下,胞浆内的细胞质模式识别受体(pattern recognition receptors,PRRs)被激活[10].在激活的PRRs的刺激下,caspase-1前体可被含有caspase募集结构域(caspase recruitment domain,CARD)的PRRs直接募集或经含CARD结构的凋亡相关斑点样蛋白(apoptosis-associated speck-like protein containing a CARD,ASC)间接募集,共同组装为caspase-1依赖性炎性小体.随后,该小体可通过自裂激活为caspase-1.而后,caspase-1以如下2种方式引起细胞焦亡:①将Gasdermin蛋白D(Gasdermin-D,GSDMD)裂解为N-GSDMD.后者能穿透细胞膜并在膜上形成孔隙,引起细胞渗透压改变,从而导致细胞肿胀至破裂.②把白细胞介素-1β(interleukin-1β,IL-1β)、IL-18的前体裂解为成熟的IL-1β、IL-18,引发炎症反应[11]. ...
... 在上述过程中,PRRs是细胞焦亡启动的关键.作为PRRs的主要组成部分,NOD样受体(NOD-like receptors,NLRs)也在细胞焦亡的启动中扮演了重要角色.常见的NLRs有NOD样受体热蛋白结构域相关蛋白1(NOD-like receptor thermal protein domain associated protein 1,NLRP1)、NLRP3、含caspase激活和募集结构域的NOD样受体4(NOD-like receptor containing a caspase activating and recruitment domain 4,NLRC4)、NLRP6和NLRP9b,它们均以不同的方式参与了细胞焦亡的启动[12]:①NLRP1通过特异性识别炭疽芽孢杆菌的致死因子蛋白酶、福氏志贺菌分泌的泛素连接酶等,激活caspase-1后启动细胞焦亡[13].②NLRP3在受到微生物、颗粒物质(如错误折叠的蛋白质)和损伤相关分子(如细胞外ATP)等的激活后,可诱导细胞焦亡的发生[14],如细胞外ATP通过激活P2X嘌呤受体7(P2X7)引起K+外排,进而激活NLRP3并促使IL-1β分泌及caspase-1活化,启动细胞焦亡的依赖caspase-1途径[11].③NLRC4炎性小体的NLR家族凋亡抑制蛋白(NLR family apoptosis inhibitory protein,NAIP)结构在特异性识别PAMPs后,可形成NLRC4炎性小体复合物,该复合物通过激活caspase-1来启动细胞焦亡.④NLRP6和NLRP9b主要在肠上皮细胞中表达,与结肠炎的进展相关[15].前者可通过激活caspase-1来裂解GSDMD,导致细胞焦亡发生;后者既可通过caspase-1裂解GSDMD引发细胞焦亡,又可通过caspase-1将IL-18前体裂解为IL-18来引起炎症反应,最终激活细胞焦亡的经典途径. ...
1
... 在上述过程中,PRRs是细胞焦亡启动的关键.作为PRRs的主要组成部分,NOD样受体(NOD-like receptors,NLRs)也在细胞焦亡的启动中扮演了重要角色.常见的NLRs有NOD样受体热蛋白结构域相关蛋白1(NOD-like receptor thermal protein domain associated protein 1,NLRP1)、NLRP3、含caspase激活和募集结构域的NOD样受体4(NOD-like receptor containing a caspase activating and recruitment domain 4,NLRC4)、NLRP6和NLRP9b,它们均以不同的方式参与了细胞焦亡的启动[12]:①NLRP1通过特异性识别炭疽芽孢杆菌的致死因子蛋白酶、福氏志贺菌分泌的泛素连接酶等,激活caspase-1后启动细胞焦亡[13].②NLRP3在受到微生物、颗粒物质(如错误折叠的蛋白质)和损伤相关分子(如细胞外ATP)等的激活后,可诱导细胞焦亡的发生[14],如细胞外ATP通过激活P2X嘌呤受体7(P2X7)引起K+外排,进而激活NLRP3并促使IL-1β分泌及caspase-1活化,启动细胞焦亡的依赖caspase-1途径[11].③NLRC4炎性小体的NLR家族凋亡抑制蛋白(NLR family apoptosis inhibitory protein,NAIP)结构在特异性识别PAMPs后,可形成NLRC4炎性小体复合物,该复合物通过激活caspase-1来启动细胞焦亡.④NLRP6和NLRP9b主要在肠上皮细胞中表达,与结肠炎的进展相关[15].前者可通过激活caspase-1来裂解GSDMD,导致细胞焦亡发生;后者既可通过caspase-1裂解GSDMD引发细胞焦亡,又可通过caspase-1将IL-18前体裂解为IL-18来引起炎症反应,最终激活细胞焦亡的经典途径. ...
1
... 在上述过程中,PRRs是细胞焦亡启动的关键.作为PRRs的主要组成部分,NOD样受体(NOD-like receptors,NLRs)也在细胞焦亡的启动中扮演了重要角色.常见的NLRs有NOD样受体热蛋白结构域相关蛋白1(NOD-like receptor thermal protein domain associated protein 1,NLRP1)、NLRP3、含caspase激活和募集结构域的NOD样受体4(NOD-like receptor containing a caspase activating and recruitment domain 4,NLRC4)、NLRP6和NLRP9b,它们均以不同的方式参与了细胞焦亡的启动[12]:①NLRP1通过特异性识别炭疽芽孢杆菌的致死因子蛋白酶、福氏志贺菌分泌的泛素连接酶等,激活caspase-1后启动细胞焦亡[13].②NLRP3在受到微生物、颗粒物质(如错误折叠的蛋白质)和损伤相关分子(如细胞外ATP)等的激活后,可诱导细胞焦亡的发生[14],如细胞外ATP通过激活P2X嘌呤受体7(P2X7)引起K+外排,进而激活NLRP3并促使IL-1β分泌及caspase-1活化,启动细胞焦亡的依赖caspase-1途径[11].③NLRC4炎性小体的NLR家族凋亡抑制蛋白(NLR family apoptosis inhibitory protein,NAIP)结构在特异性识别PAMPs后,可形成NLRC4炎性小体复合物,该复合物通过激活caspase-1来启动细胞焦亡.④NLRP6和NLRP9b主要在肠上皮细胞中表达,与结肠炎的进展相关[15].前者可通过激活caspase-1来裂解GSDMD,导致细胞焦亡发生;后者既可通过caspase-1裂解GSDMD引发细胞焦亡,又可通过caspase-1将IL-18前体裂解为IL-18来引起炎症反应,最终激活细胞焦亡的经典途径. ...
1
... 在上述过程中,PRRs是细胞焦亡启动的关键.作为PRRs的主要组成部分,NOD样受体(NOD-like receptors,NLRs)也在细胞焦亡的启动中扮演了重要角色.常见的NLRs有NOD样受体热蛋白结构域相关蛋白1(NOD-like receptor thermal protein domain associated protein 1,NLRP1)、NLRP3、含caspase激活和募集结构域的NOD样受体4(NOD-like receptor containing a caspase activating and recruitment domain 4,NLRC4)、NLRP6和NLRP9b,它们均以不同的方式参与了细胞焦亡的启动[12]:①NLRP1通过特异性识别炭疽芽孢杆菌的致死因子蛋白酶、福氏志贺菌分泌的泛素连接酶等,激活caspase-1后启动细胞焦亡[13].②NLRP3在受到微生物、颗粒物质(如错误折叠的蛋白质)和损伤相关分子(如细胞外ATP)等的激活后,可诱导细胞焦亡的发生[14],如细胞外ATP通过激活P2X嘌呤受体7(P2X7)引起K+外排,进而激活NLRP3并促使IL-1β分泌及caspase-1活化,启动细胞焦亡的依赖caspase-1途径[11].③NLRC4炎性小体的NLR家族凋亡抑制蛋白(NLR family apoptosis inhibitory protein,NAIP)结构在特异性识别PAMPs后,可形成NLRC4炎性小体复合物,该复合物通过激活caspase-1来启动细胞焦亡.④NLRP6和NLRP9b主要在肠上皮细胞中表达,与结肠炎的进展相关[15].前者可通过激活caspase-1来裂解GSDMD,导致细胞焦亡发生;后者既可通过caspase-1裂解GSDMD引发细胞焦亡,又可通过caspase-1将IL-18前体裂解为IL-18来引起炎症反应,最终激活细胞焦亡的经典途径. ...
1
... 在上述过程中,PRRs是细胞焦亡启动的关键.作为PRRs的主要组成部分,NOD样受体(NOD-like receptors,NLRs)也在细胞焦亡的启动中扮演了重要角色.常见的NLRs有NOD样受体热蛋白结构域相关蛋白1(NOD-like receptor thermal protein domain associated protein 1,NLRP1)、NLRP3、含caspase激活和募集结构域的NOD样受体4(NOD-like receptor containing a caspase activating and recruitment domain 4,NLRC4)、NLRP6和NLRP9b,它们均以不同的方式参与了细胞焦亡的启动[12]:①NLRP1通过特异性识别炭疽芽孢杆菌的致死因子蛋白酶、福氏志贺菌分泌的泛素连接酶等,激活caspase-1后启动细胞焦亡[13].②NLRP3在受到微生物、颗粒物质(如错误折叠的蛋白质)和损伤相关分子(如细胞外ATP)等的激活后,可诱导细胞焦亡的发生[14],如细胞外ATP通过激活P2X嘌呤受体7(P2X7)引起K+外排,进而激活NLRP3并促使IL-1β分泌及caspase-1活化,启动细胞焦亡的依赖caspase-1途径[11].③NLRC4炎性小体的NLR家族凋亡抑制蛋白(NLR family apoptosis inhibitory protein,NAIP)结构在特异性识别PAMPs后,可形成NLRC4炎性小体复合物,该复合物通过激活caspase-1来启动细胞焦亡.④NLRP6和NLRP9b主要在肠上皮细胞中表达,与结肠炎的进展相关[15].前者可通过激活caspase-1来裂解GSDMD,导致细胞焦亡发生;后者既可通过caspase-1裂解GSDMD引发细胞焦亡,又可通过caspase-1将IL-18前体裂解为IL-18来引起炎症反应,最终激活细胞焦亡的经典途径. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}