Research progress in the roles of Notch signaling pathway during fracture healing
GUO Liqiang,, ZHAO Shitian, SHU Bing,
Longhua Hospital, Shanghai University of Traditional Chinese Medicine; Spine Institute, Shanghai Academy of Traditional Chinese Medicine; Key Laboratory of Ministry of Education for Theory and Treatment of Bones and Muscles, Shanghai 200032, China
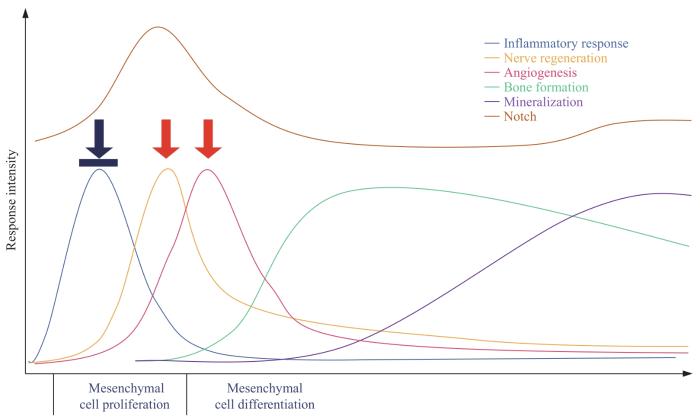
Fracture is the most common large-organ injury in humans. It takes several months or even longer from the onset to the completion of bone reconstruction. In severe cases, delayed healing or even bone discontinuity can occur. The current treatment of fractures mainly pursues the completion of clinical healing and bone healing, and the healing process is divided into three stages: early, intermediate and late. Various factors affect the speed of the healing process, among which signaling pathways and cytokines play an important role in fracture healing, so understanding the important role of signaling pathways and cytokines is important for treating fractures and promoting fracture healing. Recent studies have shown that the Notch signaling pathway can affect cell proliferation and differentiation, inflammatory response, bone reconstruction, angiogenesis and nerve regeneration during fracture healing, and its changes are also closely related to mechanical stimulation and other factors. Therefore, this paper reviews the research progress in the role of Notch signaling pathway in various aspects of fracture healing from the perspectives of cell proliferation and differentiation, inflammatory response, bone reconstruction, angiogenesis, nerve regeneration and mechanical stimulation, and provides new research directions and therapeutic strategies for fracture healing treatment.
GUO Liqiang, ZHAO Shitian, SHU Bing. Research progress in the roles of Notch signaling pathway during fracture healing. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(2): 222-229 doi:10.3969/j.issn.1674-8115.2023.02.012
利用Osx-cre工具小鼠,在成骨前体细胞中敲除Notch1和Notch2,可以导致小鼠出现短暂的成骨细胞数量、骨形成和松质骨体积的增加,同时伴有骨吸收活性下降[26]。一致的是,利用Ⅰ型胶原启动子在成骨细胞中过表达NICD,则会使小鼠出现骨小梁数量、成骨细胞数量和骨体积降低的表型[27]。体外实验[28]中,在人骨髓MSC中分别过表达NICD和Jagged1,碱性磷酸酶(alkaline phosphatase,ALP)活性明显增加,提示Notch信号通路的激活可以显著促进骨髓MSC的成骨分化。但ZANOTTI等[27]发现,在小鼠原代骨髓MSC和成骨细胞中过表达NICD,会导致ALP表达降低,骨形成能力下降;同时该研究还提出,Notch信号过度激活导致的成骨细胞骨形成能力下降,可能与抑制无翅型MMTV整合位点家族(wingless-type MMTV integration site family,Wnt)/β-catenin信号通路有关。
JI等[29]则发现,在骨髓MSC成骨分化的终末期和被矿化基质包埋的骨细胞中,Notch信号也是激活的。抑制Notch通路,可以显著抑制矿化相关基因的表达,降低骨髓MSC的矿化能力。体内研究也表明,间断性注射Notch通路激活剂,小鼠、大鼠和去卵巢大鼠的骨形成均明显增加,而持续注射Notch通路激活剂则对骨形成没有显著作用。因此,他们认为在骨形成的过程中,Notch信号通路的激活呈现了双阶段的特点:即分别在成骨细胞分化的早期和晚期激活,分别促进了前成骨细胞的增殖和成熟成骨细胞向骨细胞分化。Notch信号通路对破骨细胞形成和骨吸收也有一定的调控作用。在小鼠的破骨细胞前体细胞中激活Notch信号通路的转录因子RBPJ可以抑制TNF-α诱导的破骨细胞形成和骨吸收,其机制主要是通过抑制细胞原癌基因(immediate early gene,c-fos)和B淋巴细胞诱导成熟蛋白-1的活化,抑制破骨细胞生成的主调节因子活化T细胞的核因子-1(nuclear factor of activated T cells 1,NFATc-1)的生成,从而阻止破骨细胞形成抑制因子的下调[30]。与RBPJ对破骨细胞生成的抑制作用相反,在破骨细胞前体细胞持续激活Notch2,则细胞对核因子κB激活性受体配体(receptor activator for nuclear factor-κB,RANKL)诱导的破骨细胞生成和骨吸收反应更强;与之一致的是,在破骨细胞前体细胞持续激活Notch2,小鼠也会出现破骨细胞数量上升,松质骨和皮质骨骨量下降的表型[31]。GOEL等[32]在体外抑制小鼠破骨细胞前体细胞中的Notch信号,发现该细胞虽然不能有效融合,但ALP、组织蛋白酶K(cathepsin K)、基质金属蛋白酶9(matrix metalloproteinase 9,MMP9)等破骨标志物表达水平并无明显变化,表明早期破骨细胞的分化并不需要Notch信号的参与,Notch信号可能影响破骨细胞的成熟。
体内研究[44]表明,小鼠后肢胫骨骨折后施加相应机械刺激,Notch信号通路相关分子的基因表达均上调,其骨痂组织中Notch下游基因Hes family BHLH transcription factor 1(Hes1)、Hes related family BHLH transcription factor with YRPW motif 1(Hey1)和Hey2的表达增加了6倍,骨折处MSC的增殖和分化显著增强,骨愈合进程明显加快。此外,体外实验[45]证明,力学因素可以显著提高体外培养的骨髓MSC中Notch信号通路的相关基因,包括Notch受体Notch1和Notch2和下游的Hes1、Hey1、Hey2等的表达。同样,给予前成骨细胞间断压力刺激,可以使细胞中Notch下游靶基因Hes1、Hey1的表达上调,且该作用是通过TGF-β通路介导的。目前虽已证实Notch在成骨细胞谱系中的表达受到力学因素的调节,但力学刺激可以导致硬化蛋白(sclerostin,Sost)基因水平降低,Notch信号的激活可以逆转其抑制作用,上调Sost的基因表达水平,后者则是Wnt/β-catenin信号通路的抑制因子[46]。尽管力学刺激下的Notch信号会促进Sost的表达,但因其具有促进细胞增殖的作用,可以储备大量的MSC或成骨前体细胞,为后续促进骨形成奠定基础。因此,Notch信号从高表达向低表达转换,是启动骨形成的重要节点。
The design of the paper, and manuscript drafting and revision were conducted by SHU Bing, who is the corresponding author of this paper. The literature reading and manuscript drafting were conducted by GUO Liqiang and ZHAO Shitian. All authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
SHAYA O, BINSHTOK U, HERSCH M, et al. Cell-cell contact area affects Notch signaling and Notch-dependent patterning[J]. Dev Cell, 2017, 40(5): 505-511.e6.
FAYYAZ S, ATTAR R, XU B J, et al. Realizing the potential of blueberry as natural inhibitor of metastasis and powerful apoptosis inducer: tapping the treasure trove for effective regulation of cell signaling pathways[J]. Anticancer Agents Med Chem, 2020, 20(15): 1780-1786.
DISHOWITZ M I, MUTYABA P L, TAKACS J D, et al. Systemic inhibition of canonical Notch signaling results in sustained callus inflammation and alters multiple phases of fracture healing[J]. PLoS One, 2013, 8(7): e68726.
KEEWAN E, NASER S A. The role of Notch signaling in macrophages during inflammation and infection: implication in rheumatoid arthritis?[J]. Cells, 2020, 9(1): 111.
HILTON M J, TU X L, WU X M, et al. Notch signaling maintains bone marrow mesenchymal progenitors by suppressing osteoblast differentiation[J]. Nat Med, 2008, 14(3): 306-314.
ZHANG Q H, WANG C M, LIU Z L, et al. Notch signal suppresses toll-like receptor-triggered inflammatory responses in macrophages by inhibiting extracellular signal-regulated kinase 1/2-mediated nuclear factor κB activation[J]. J Biol Chem, 2012, 287(9): 6208-6217.
HALL S R R, JIANG Y J, LEARY E, et al. Identification and isolation of small CD44-negative mesenchymal stem/progenitor cells from human bone marrow using elutriation and polychromatic flow cytometry[J]. Stem Cells Transl Med, 2013, 2(8): 567-578.
SONG K, HUANG M Q, SHI Q, et al. Cultivation and identification of rat bone marrow-derived mesenchymal stem cells[J]. Mol Med Rep, 2014, 10(2): 755-760.
DISHOWITZ M I, TERKHORN S P, BOSTIC S A, et al. Notch signaling components are upregulated during both endochondral and intramembranous bone regeneration[J]. J Orthop Res, 2012, 30(2): 296-303.
MATTHEWS B G, GRCEVIC D, WANG L P, et al. Analysis of αSMA-labeled progenitor cell commitment identifies Notch signaling as an important pathway in fracture healing[J]. J Bone Miner Res, 2014, 29(5): 1283-1294.
WANG C, INZANA J A, MIRANDO A J, et al. NOTCH signaling in skeletal progenitors is critical for fracture repair[J]. J Clin Invest, 2016, 126(4): 1471-1481.
SEMENOVA D, BOGDANOVA M, KOSTINA A, et al. Dose-dependent mechanism of Notch action in promoting osteogenic differentiation of mesenchymal stem cells[J]. Cell Tissue Res, 2020, 379(1): 169-179.
UGARTE F, RYSER M, THIEME S, et al. Notch signaling enhances osteogenic differentiation while inhibiting adipogenesis in primary human bone marrow stromal cells[J]. Exp Hematol, 2009, 37(7): 867-875.e1.
ZHAO B H, GRIMES S N, LI S S, et al. TNF-induced osteoclastogenesis and inflammatory bone resorption are inhibited by transcription factor RBP-J[J]. J Exp Med, 2012, 209(2): 319-334.
GOEL P N, MOHARRER Y, HEBB J H, et al. Suppression of Notch signaling in osteoclasts improves bone regeneration and healing[J]. J Orthop Res, 2019, 37(10): 2089-2103.
SAHARA M, HANSSON E M, WERNET O, et al. Manipulation of a VEGF-Notch signaling circuit drives formation of functional vascular endothelial progenitors from human pluripotent stem cells[J]. Cell Res, 2015, 25(1): 148.
KUSUMBE A P, RAMASAMY S K, ADAMS R H. Coupling of angiogenesis and osteogenesis by a specific vessel subtype in bone[J]. Nature, 2014, 507(7492): 323-328.
RAMASAMY S K, KUSUMBE A P, WANG L, et al. Endothelial Notch activity promotes angiogenesis and osteogenesis in bone[J]. Nature, 2014, 507(7492): 376-380.
YANG M, LI C J, SUN X, et al. MiR-497~195 cluster regulates angiogenesis during coupling with osteogenesis by maintaining endothelial Notch and HIF-1α activity[J]. Nat Commun, 2017, 8: 16003.
ZHU Y, RUAN Z, LIN Z Y, et al. The association between CD31hiEmcnhi endothelial cells and bone mineral density in Chinese women[J]. J Bone Miner Metab, 2019, 37(6): 987-995.
PFLANZ D, BIRKHOLD A I, ALBIOL L, et al. Sost deficiency led to a greater cortical bone formation response to mechanical loading and altered gene expression[J]. Sci Rep, 2017, 7(1): 9435.
ZIOUTI F, EBERT R, RUMMLER M, et al. NOTCH signaling is activated through mechanical strain in human bone marrow-derived mesenchymal stromal cells[J]. Stem Cells Int, 2019, 2019: 5150634.
NIEDERMAIR T, STRAUB R H, BROCHHAUSEN C, et al. Impact of the sensory and sympathetic nervous system on fracture healing in ovariectomized mice[J]. Int J Mol Sci, 2020, 21(2): 405.
WANG J, REN K Y, WANG Y H, et al. Effect of active Notch signaling system on the early repair of rat sciatic nerve injury[J]. Artif Cells Nanomed Biotechnol, 2015, 43(6): 383-389.
KAMIŃSKA A, MAREK S, PARDYAK L, et al. Crosstalk between androgen-ZIP9 signaling and Notch pathway in rodent Sertoli cells[J]. Int J Mol Sci, 2020, 21(21): 8275.
... 利用Osx-cre工具小鼠,在成骨前体细胞中敲除Notch1和Notch2,可以导致小鼠出现短暂的成骨细胞数量、骨形成和松质骨体积的增加,同时伴有骨吸收活性下降[26].一致的是,利用Ⅰ型胶原启动子在成骨细胞中过表达NICD,则会使小鼠出现骨小梁数量、成骨细胞数量和骨体积降低的表型[27].体外实验[28]中,在人骨髓MSC中分别过表达NICD和Jagged1,碱性磷酸酶(alkaline phosphatase,ALP)活性明显增加,提示Notch信号通路的激活可以显著促进骨髓MSC的成骨分化.但ZANOTTI等[27]发现,在小鼠原代骨髓MSC和成骨细胞中过表达NICD,会导致ALP表达降低,骨形成能力下降;同时该研究还提出,Notch信号过度激活导致的成骨细胞骨形成能力下降,可能与抑制无翅型MMTV整合位点家族(wingless-type MMTV integration site family,Wnt)/β-catenin信号通路有关. ...
2
... 利用Osx-cre工具小鼠,在成骨前体细胞中敲除Notch1和Notch2,可以导致小鼠出现短暂的成骨细胞数量、骨形成和松质骨体积的增加,同时伴有骨吸收活性下降[26].一致的是,利用Ⅰ型胶原启动子在成骨细胞中过表达NICD,则会使小鼠出现骨小梁数量、成骨细胞数量和骨体积降低的表型[27].体外实验[28]中,在人骨髓MSC中分别过表达NICD和Jagged1,碱性磷酸酶(alkaline phosphatase,ALP)活性明显增加,提示Notch信号通路的激活可以显著促进骨髓MSC的成骨分化.但ZANOTTI等[27]发现,在小鼠原代骨髓MSC和成骨细胞中过表达NICD,会导致ALP表达降低,骨形成能力下降;同时该研究还提出,Notch信号过度激活导致的成骨细胞骨形成能力下降,可能与抑制无翅型MMTV整合位点家族(wingless-type MMTV integration site family,Wnt)/β-catenin信号通路有关. ...
... [27]发现,在小鼠原代骨髓MSC和成骨细胞中过表达NICD,会导致ALP表达降低,骨形成能力下降;同时该研究还提出,Notch信号过度激活导致的成骨细胞骨形成能力下降,可能与抑制无翅型MMTV整合位点家族(wingless-type MMTV integration site family,Wnt)/β-catenin信号通路有关. ...
1
... 利用Osx-cre工具小鼠,在成骨前体细胞中敲除Notch1和Notch2,可以导致小鼠出现短暂的成骨细胞数量、骨形成和松质骨体积的增加,同时伴有骨吸收活性下降[26].一致的是,利用Ⅰ型胶原启动子在成骨细胞中过表达NICD,则会使小鼠出现骨小梁数量、成骨细胞数量和骨体积降低的表型[27].体外实验[28]中,在人骨髓MSC中分别过表达NICD和Jagged1,碱性磷酸酶(alkaline phosphatase,ALP)活性明显增加,提示Notch信号通路的激活可以显著促进骨髓MSC的成骨分化.但ZANOTTI等[27]发现,在小鼠原代骨髓MSC和成骨细胞中过表达NICD,会导致ALP表达降低,骨形成能力下降;同时该研究还提出,Notch信号过度激活导致的成骨细胞骨形成能力下降,可能与抑制无翅型MMTV整合位点家族(wingless-type MMTV integration site family,Wnt)/β-catenin信号通路有关. ...
1
... JI等[29]则发现,在骨髓MSC成骨分化的终末期和被矿化基质包埋的骨细胞中,Notch信号也是激活的.抑制Notch通路,可以显著抑制矿化相关基因的表达,降低骨髓MSC的矿化能力.体内研究也表明,间断性注射Notch通路激活剂,小鼠、大鼠和去卵巢大鼠的骨形成均明显增加,而持续注射Notch通路激活剂则对骨形成没有显著作用.因此,他们认为在骨形成的过程中,Notch信号通路的激活呈现了双阶段的特点:即分别在成骨细胞分化的早期和晚期激活,分别促进了前成骨细胞的增殖和成熟成骨细胞向骨细胞分化.Notch信号通路对破骨细胞形成和骨吸收也有一定的调控作用.在小鼠的破骨细胞前体细胞中激活Notch信号通路的转录因子RBPJ可以抑制TNF-α诱导的破骨细胞形成和骨吸收,其机制主要是通过抑制细胞原癌基因(immediate early gene,c-fos)和B淋巴细胞诱导成熟蛋白-1的活化,抑制破骨细胞生成的主调节因子活化T细胞的核因子-1(nuclear factor of activated T cells 1,NFATc-1)的生成,从而阻止破骨细胞形成抑制因子的下调[30].与RBPJ对破骨细胞生成的抑制作用相反,在破骨细胞前体细胞持续激活Notch2,则细胞对核因子κB激活性受体配体(receptor activator for nuclear factor-κB,RANKL)诱导的破骨细胞生成和骨吸收反应更强;与之一致的是,在破骨细胞前体细胞持续激活Notch2,小鼠也会出现破骨细胞数量上升,松质骨和皮质骨骨量下降的表型[31].GOEL等[32]在体外抑制小鼠破骨细胞前体细胞中的Notch信号,发现该细胞虽然不能有效融合,但ALP、组织蛋白酶K(cathepsin K)、基质金属蛋白酶9(matrix metalloproteinase 9,MMP9)等破骨标志物表达水平并无明显变化,表明早期破骨细胞的分化并不需要Notch信号的参与,Notch信号可能影响破骨细胞的成熟. ...
1
... JI等[29]则发现,在骨髓MSC成骨分化的终末期和被矿化基质包埋的骨细胞中,Notch信号也是激活的.抑制Notch通路,可以显著抑制矿化相关基因的表达,降低骨髓MSC的矿化能力.体内研究也表明,间断性注射Notch通路激活剂,小鼠、大鼠和去卵巢大鼠的骨形成均明显增加,而持续注射Notch通路激活剂则对骨形成没有显著作用.因此,他们认为在骨形成的过程中,Notch信号通路的激活呈现了双阶段的特点:即分别在成骨细胞分化的早期和晚期激活,分别促进了前成骨细胞的增殖和成熟成骨细胞向骨细胞分化.Notch信号通路对破骨细胞形成和骨吸收也有一定的调控作用.在小鼠的破骨细胞前体细胞中激活Notch信号通路的转录因子RBPJ可以抑制TNF-α诱导的破骨细胞形成和骨吸收,其机制主要是通过抑制细胞原癌基因(immediate early gene,c-fos)和B淋巴细胞诱导成熟蛋白-1的活化,抑制破骨细胞生成的主调节因子活化T细胞的核因子-1(nuclear factor of activated T cells 1,NFATc-1)的生成,从而阻止破骨细胞形成抑制因子的下调[30].与RBPJ对破骨细胞生成的抑制作用相反,在破骨细胞前体细胞持续激活Notch2,则细胞对核因子κB激活性受体配体(receptor activator for nuclear factor-κB,RANKL)诱导的破骨细胞生成和骨吸收反应更强;与之一致的是,在破骨细胞前体细胞持续激活Notch2,小鼠也会出现破骨细胞数量上升,松质骨和皮质骨骨量下降的表型[31].GOEL等[32]在体外抑制小鼠破骨细胞前体细胞中的Notch信号,发现该细胞虽然不能有效融合,但ALP、组织蛋白酶K(cathepsin K)、基质金属蛋白酶9(matrix metalloproteinase 9,MMP9)等破骨标志物表达水平并无明显变化,表明早期破骨细胞的分化并不需要Notch信号的参与,Notch信号可能影响破骨细胞的成熟. ...
1
... JI等[29]则发现,在骨髓MSC成骨分化的终末期和被矿化基质包埋的骨细胞中,Notch信号也是激活的.抑制Notch通路,可以显著抑制矿化相关基因的表达,降低骨髓MSC的矿化能力.体内研究也表明,间断性注射Notch通路激活剂,小鼠、大鼠和去卵巢大鼠的骨形成均明显增加,而持续注射Notch通路激活剂则对骨形成没有显著作用.因此,他们认为在骨形成的过程中,Notch信号通路的激活呈现了双阶段的特点:即分别在成骨细胞分化的早期和晚期激活,分别促进了前成骨细胞的增殖和成熟成骨细胞向骨细胞分化.Notch信号通路对破骨细胞形成和骨吸收也有一定的调控作用.在小鼠的破骨细胞前体细胞中激活Notch信号通路的转录因子RBPJ可以抑制TNF-α诱导的破骨细胞形成和骨吸收,其机制主要是通过抑制细胞原癌基因(immediate early gene,c-fos)和B淋巴细胞诱导成熟蛋白-1的活化,抑制破骨细胞生成的主调节因子活化T细胞的核因子-1(nuclear factor of activated T cells 1,NFATc-1)的生成,从而阻止破骨细胞形成抑制因子的下调[30].与RBPJ对破骨细胞生成的抑制作用相反,在破骨细胞前体细胞持续激活Notch2,则细胞对核因子κB激活性受体配体(receptor activator for nuclear factor-κB,RANKL)诱导的破骨细胞生成和骨吸收反应更强;与之一致的是,在破骨细胞前体细胞持续激活Notch2,小鼠也会出现破骨细胞数量上升,松质骨和皮质骨骨量下降的表型[31].GOEL等[32]在体外抑制小鼠破骨细胞前体细胞中的Notch信号,发现该细胞虽然不能有效融合,但ALP、组织蛋白酶K(cathepsin K)、基质金属蛋白酶9(matrix metalloproteinase 9,MMP9)等破骨标志物表达水平并无明显变化,表明早期破骨细胞的分化并不需要Notch信号的参与,Notch信号可能影响破骨细胞的成熟. ...
1
... JI等[29]则发现,在骨髓MSC成骨分化的终末期和被矿化基质包埋的骨细胞中,Notch信号也是激活的.抑制Notch通路,可以显著抑制矿化相关基因的表达,降低骨髓MSC的矿化能力.体内研究也表明,间断性注射Notch通路激活剂,小鼠、大鼠和去卵巢大鼠的骨形成均明显增加,而持续注射Notch通路激活剂则对骨形成没有显著作用.因此,他们认为在骨形成的过程中,Notch信号通路的激活呈现了双阶段的特点:即分别在成骨细胞分化的早期和晚期激活,分别促进了前成骨细胞的增殖和成熟成骨细胞向骨细胞分化.Notch信号通路对破骨细胞形成和骨吸收也有一定的调控作用.在小鼠的破骨细胞前体细胞中激活Notch信号通路的转录因子RBPJ可以抑制TNF-α诱导的破骨细胞形成和骨吸收,其机制主要是通过抑制细胞原癌基因(immediate early gene,c-fos)和B淋巴细胞诱导成熟蛋白-1的活化,抑制破骨细胞生成的主调节因子活化T细胞的核因子-1(nuclear factor of activated T cells 1,NFATc-1)的生成,从而阻止破骨细胞形成抑制因子的下调[30].与RBPJ对破骨细胞生成的抑制作用相反,在破骨细胞前体细胞持续激活Notch2,则细胞对核因子κB激活性受体配体(receptor activator for nuclear factor-κB,RANKL)诱导的破骨细胞生成和骨吸收反应更强;与之一致的是,在破骨细胞前体细胞持续激活Notch2,小鼠也会出现破骨细胞数量上升,松质骨和皮质骨骨量下降的表型[31].GOEL等[32]在体外抑制小鼠破骨细胞前体细胞中的Notch信号,发现该细胞虽然不能有效融合,但ALP、组织蛋白酶K(cathepsin K)、基质金属蛋白酶9(matrix metalloproteinase 9,MMP9)等破骨标志物表达水平并无明显变化,表明早期破骨细胞的分化并不需要Notch信号的参与,Notch信号可能影响破骨细胞的成熟. ...
... 体内研究[44]表明,小鼠后肢胫骨骨折后施加相应机械刺激,Notch信号通路相关分子的基因表达均上调,其骨痂组织中Notch下游基因Hes family BHLH transcription factor 1(Hes1)、Hes related family BHLH transcription factor with YRPW motif 1(Hey1)和Hey2的表达增加了6倍,骨折处MSC的增殖和分化显著增强,骨愈合进程明显加快.此外,体外实验[45]证明,力学因素可以显著提高体外培养的骨髓MSC中Notch信号通路的相关基因,包括Notch受体Notch1和Notch2和下游的Hes1、Hey1、Hey2等的表达.同样,给予前成骨细胞间断压力刺激,可以使细胞中Notch下游靶基因Hes1、Hey1的表达上调,且该作用是通过TGF-β通路介导的.目前虽已证实Notch在成骨细胞谱系中的表达受到力学因素的调节,但力学刺激可以导致硬化蛋白(sclerostin,Sost)基因水平降低,Notch信号的激活可以逆转其抑制作用,上调Sost的基因表达水平,后者则是Wnt/β-catenin信号通路的抑制因子[46].尽管力学刺激下的Notch信号会促进Sost的表达,但因其具有促进细胞增殖的作用,可以储备大量的MSC或成骨前体细胞,为后续促进骨形成奠定基础.因此,Notch信号从高表达向低表达转换,是启动骨形成的重要节点. ...
1
... 体内研究[44]表明,小鼠后肢胫骨骨折后施加相应机械刺激,Notch信号通路相关分子的基因表达均上调,其骨痂组织中Notch下游基因Hes family BHLH transcription factor 1(Hes1)、Hes related family BHLH transcription factor with YRPW motif 1(Hey1)和Hey2的表达增加了6倍,骨折处MSC的增殖和分化显著增强,骨愈合进程明显加快.此外,体外实验[45]证明,力学因素可以显著提高体外培养的骨髓MSC中Notch信号通路的相关基因,包括Notch受体Notch1和Notch2和下游的Hes1、Hey1、Hey2等的表达.同样,给予前成骨细胞间断压力刺激,可以使细胞中Notch下游靶基因Hes1、Hey1的表达上调,且该作用是通过TGF-β通路介导的.目前虽已证实Notch在成骨细胞谱系中的表达受到力学因素的调节,但力学刺激可以导致硬化蛋白(sclerostin,Sost)基因水平降低,Notch信号的激活可以逆转其抑制作用,上调Sost的基因表达水平,后者则是Wnt/β-catenin信号通路的抑制因子[46].尽管力学刺激下的Notch信号会促进Sost的表达,但因其具有促进细胞增殖的作用,可以储备大量的MSC或成骨前体细胞,为后续促进骨形成奠定基础.因此,Notch信号从高表达向低表达转换,是启动骨形成的重要节点. ...
1
... 体内研究[44]表明,小鼠后肢胫骨骨折后施加相应机械刺激,Notch信号通路相关分子的基因表达均上调,其骨痂组织中Notch下游基因Hes family BHLH transcription factor 1(Hes1)、Hes related family BHLH transcription factor with YRPW motif 1(Hey1)和Hey2的表达增加了6倍,骨折处MSC的增殖和分化显著增强,骨愈合进程明显加快.此外,体外实验[45]证明,力学因素可以显著提高体外培养的骨髓MSC中Notch信号通路的相关基因,包括Notch受体Notch1和Notch2和下游的Hes1、Hey1、Hey2等的表达.同样,给予前成骨细胞间断压力刺激,可以使细胞中Notch下游靶基因Hes1、Hey1的表达上调,且该作用是通过TGF-β通路介导的.目前虽已证实Notch在成骨细胞谱系中的表达受到力学因素的调节,但力学刺激可以导致硬化蛋白(sclerostin,Sost)基因水平降低,Notch信号的激活可以逆转其抑制作用,上调Sost的基因表达水平,后者则是Wnt/β-catenin信号通路的抑制因子[46].尽管力学刺激下的Notch信号会促进Sost的表达,但因其具有促进细胞增殖的作用,可以储备大量的MSC或成骨前体细胞,为后续促进骨形成奠定基础.因此,Notch信号从高表达向低表达转换,是启动骨形成的重要节点. ...



{kind=link}
{kind=link}