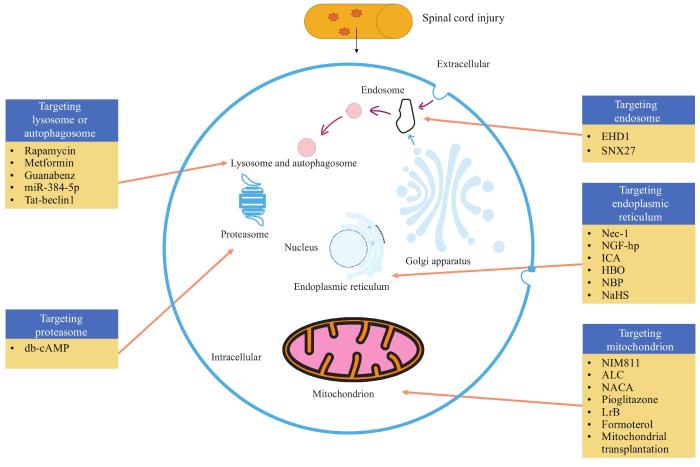
Spinal cord injury is a serious disease that greatly affects the sensory function, motor function and autonomic nervous function of patients. It not only brings serious physical and mental harm to patients but also causes huge economic burden to the society. With the development of medical technology, the research on the internal mechanism of spinal cord injury is also deepening, and the treatment methods of spinal cord injury emerge in endlessly. However, the therapeutic effect is not satisfactory, so it is urgent to further explore new therapeutic strategies and expand new therapeutic ideas for spinal cord injury. Many studies have shown that various subcellular structures are closely related to nerve regeneration and functional recovery after spinal cord injury. Therefore, targeting subcellular structures to treat spinal cord injury plays an important role in promoting nerve regeneration and repair after spinal cord injury. This targeted therapy mainly refers to targeting a variety of subcellular structures such as mitochondria, lysosomes/autophagosomes, endoplasmic reticulum, intracellular bodies and proteasomes. A variety of therapeutic strategies targeting subcellular structures have significant therapeutic effects on spinal cord injury. Among them, mitochondrial targeting or endoplasmic reticulum targeting mainly focuses on maintaining mitochondrial energy metabolism at the injury site, while endoplasmic reticulum targeting mainly focuses on inhibiting endoplasmic reticulum stress. This article reviews the research progress of subcellular structure-targeted therapy in spinal cord injury, which is expected to be a new targeted therapy strategy for spinal cord injury and provide a new idea for the treatment of spinal cord injury.
LIU Tiexin, LIN Junqing, ZHENG Xianyou. Research progress of subcellular structure-targeted therapy in spinal cord injury. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(2): 230-236 doi:10.3969/j.issn.1674-8115.2023.02.013
溶酶体作为一种动态细胞器,参与细胞内钙信号、损伤信号的转导及细胞器间的相互作用[9],其能通过自噬、内吞和吞噬途径降解和回收生物大分子。自噬体可以通过与溶酶体融合来降解生物大分子,从而构成细胞回收机制的关键组成部分[10],其融合与脊髓损伤后神经再生高度相关[11]。溶酶体和自噬体通过作用关键信号通路中的分子影响神经生长和再生。其中研究较多的信号通路是PI3K-mTOR通路。mTOR(mammalian target of rapamycin,哺乳动物雷帕霉素靶蛋白)是一种位于溶酶体外表面的丝氨酸/苏氨酸蛋白激酶,能够影响细胞的基本生理活动,调节蛋白质的合成代谢以及自噬体生成[12-13]。
靶向溶酶体和自噬体能够通过脊髓损伤的自噬诱导发挥促神经功能恢复的作用。自噬对轴突损伤后神经再生具有促进作用。雷帕霉素,作为mTOR信号通路的抑制剂,在脊髓损伤过程中显示出对自噬的促进作用,能够减轻脊髓损伤后的组织学和功能学损伤[34]。二甲双胍作为一种降糖药物,已被证实能促进脊髓损伤后的运动功能恢复[35]。WU等[36]的研究发现二甲双胍通过促进小胶质细胞M1向M2表型极化的转化,从而极大促进髓鞘碎片清除,增强自噬小体和溶酶体融合,显示出对于脊髓损伤后自噬的诱导作用,在保护髓鞘的同时通过自噬清除损伤神经细胞中坏死的部分从而促进脊髓损伤后神经功能恢复。未折叠蛋白反应(unfolded protein response,UPR)是一种调节细胞稳态的细胞内信号[37]。有研究[38]显示,Guanabenz(一种口服活性中枢α2-肾上腺素能激动剂)通过提高UPR诱导脊髓损伤后自噬从而减少神经元死亡并促进神经功能恢复,并发现其内在机制与调节转录因子EB(transcription factor EB,TFEB)诱导自噬有关。另外,ZHOU等[39]的研究发现miR-384-5p通过调节自噬的重要启动子beclin-1的表达调节脊髓损伤后自噬,进而促进神经元存活和脊髓损伤后运动功能恢复。Tat-beclin1,作为一种特异性的自噬诱导肽,在脊髓损伤中显示出减弱损伤轴突回缩、促进神经再生和运动功能恢复的作用[40]。靶向溶酶体和自噬体治疗不仅为脊髓损伤治疗提供了新策略,还为应用现有药物进行重定位靶向治疗提供了新思路。
靶向胞内体治疗能促进脊髓损伤中胞内神经营养物质的运输。Eps15同源结构域蛋白1(Eps15 homology domain protein 1,EHD1)在胞内体系统中发挥着“看门人”的作用[50]。在WU等[51]的研究中,EHD1通过循环胞内体而显示出其对神经营养素内吞和运输的调控作用,胞内体跨膜运输也对神经突生长有着促进作用。酪氨酸激酶受体A(tyrosine kinase receptor A,TrkA),作为细胞膜上NGF的功能特异性受体,在与NGF结合时被激活。而EHD1可上调神经元TrkA的表达。该研究[51]显示通过靶向胞内体EHD1,调控TrkA的回收进而提高NGF利用效率,可达到加强对损伤神经的营养物质输送而治疗脊髓损伤的目的。分选连接蛋白27(sorting nexin family member 27,SNX27)是一种胞内体相关的介导分子,参与多种神经疾病的病理和发展过程[52],但其在脊髓损伤中的作用尚待研究。ZENG等[53]的研究发现SNX27在脊髓损伤中上调。该研究者进一步的研究发现SNX27的缺乏对脊髓损伤的神经功能学水平的恢复具有促进作用,且SNX27的缺乏可抑制神经元细胞凋亡。此外,在脊髓损伤的小鼠模型中,SNX27缺乏的小鼠神经损伤部位巨噬细胞/小胶质细胞的增殖受到抑制,并且巨噬细胞/小胶质细胞浸润和活化降低。该研究[53]提示下调SNX27是一种靶向急性神经元死亡和慢性神经炎症的潜在疗法,并有助于促进脊髓损伤后的神经组织学和功能学水平的恢复,为靶向胞内体治疗脊髓损伤的潜在靶点。
2.5 靶向蛋白酶体治疗脊髓损伤
靶向蛋白酶体治疗对脊髓损伤中的预防蛋白毒性、降低泛素化蛋白积累等起着关键作用。蛋白酶体损伤和泛素化蛋白积累与脊髓损伤中的神经退行性变相关,泛素化蛋白积累是脊髓神经元损伤时的病理特征[54]。有研究[55]发现在脊髓神经组织中升高cAMP可增加26S蛋白酶体活性。二丁基-cAMP(dibutyl-cAMP,db-cAMP)在脊髓损伤中显示出降低内源性炎症产物前列腺素J2引发的泛素化蛋白积累、蛋白酶体抑制和神经毒性的作用,可提高泛素-蛋白酶体通路(ubiquitin proteasome pathway,UPP)的各种成分水平,减轻脊髓损伤后泛素化蛋白积累和蛋白酶体损伤,从而促进脊髓损伤后神经组织学和功能学水平恢复。该研究提供了靶向蛋白酶体治疗脊髓损伤的新思路。受体相互作用蛋白激酶3(receptor interacting protein kinase 3,RIP3)是细胞坏死的关键调控因子[56]。WU等[57]的研究发现RIP3阳性神经元中蛋白酶体亚基β4型(proteasome subunit,beta type 4,PSMB4)表达显著上调,该现象提示了PSMB4可能在RIP3的调控中起作用。对PSMB4的过表达和敲低均可干预RIP3和混合谱系激酶结构域样蛋白(mixed lineage kinase domain like pseudokinase,MLKL)通路,这提示了PSMB4和RIP3与脊髓损伤后损伤神经坏死相关,可作为脊髓损伤的靶向蛋白酶体治疗潜在靶点。另外,TICA等[58]的研究通过数据分析来对脊髓损伤后持续差异表达的蛋白进行识别。该研究揭示了多个被忽视的具有生物活性和已确定具有药物功能但在脊髓损伤中表达和功能未知的治疗候选药物,其中包括上调的组织蛋白酶A、H、Z和人蛋白酶体亚基β10型(proteasome subunit beta type 10,PSMB10)等。这些蛋白酶体相关蛋白具有成为靶向蛋白酶体治疗脊髓损伤的潜在靶点的良好前景。但目前靶向蛋白酶体治疗手段仍然较少,潜在靶点众多,仍待深入研究。
The manuscript was designed, drafted and revised by LIU Tiexin. The manuscript was supervised and revised by LIN Junqing and ZHENG Xianyou. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
FEHLINGS M G, TETREAULT L A, WILSON J R, et al. A clinical practice guideline for the management of acute spinal cord injury: introduction, rationale, and scope[J]. Global Spine J, 2017, 7(3 Suppl): 84S-94S.
CHEN X Y, CHEN D, CHEN C H, et al. The epidemiology and disease burden of traumatic spinal cord injury in China: a systematic review[J]. Chinese Journal of Evidence-Based Medicine, 2018, 18(2): 143-150.
PETROVA V, NIEUWENHUIS B, FAWCETT J W, et al. Axonal organelles as molecular platforms for axon growth and regeneration after injury[J]. Int J Mol Sci, 2021, 22(4): 1798.
BARTOLÁK-SUKI E, IMSIROVIC J, NISHIBORI Y, et al. Regulation of mitochondrial structure and dynamics by the cytoskeleton and mechanical factors[J]. Int J Mol Sci, 2017, 18(8): 1812.
CAVALLUCCI V, BISICCHIA E, CENCIONI M T, et al. Acute focal brain damage alters mitochondrial dynamics and autophagy in axotomized neurons[J]. Cell Death Dis, 2014, 5(11): e1545.
MALLA B, NIESNER R, HAUSER A, et al. Imaging and analysis of neuronal mitochondria in murine acute brain slices[J]. J Neurosci Methods, 2022, 372: 109558.
SAKAMOTO K, OZAKI T, KO Y C, et al. Glycan sulfation patterns define autophagy flux at axon tip via PTPRσ-cortactin axis[J]. Nat Chem Biol, 2019, 15(7): 699-709.
KORHONEN L, LINDHOLM D. The ubiquitin proteasome system in synaptic and axonal degeneration: a new twist to an old cycle[J]. J Cell Biol, 2004, 165(1): 27-30.
LEE M, LIU Y C, CHEN C, et al. Ecm29-mediated proteasomal distribution modulates excitatory GABA responses in the developing brain[J]. J Cell Biol, 2020, 219(2): e201903033.
DI PAOLO A, GARAT J, EASTMAN G, et al. Functional genomics of axons and synapses to understand neurodegenerative diseases[J]. Front Cell Neurosci, 2021, 15: 686722.
TAKABATAKE M, GOSHIMA Y, SASAKI Y. Semaphorin-3A promotes degradation of fragile X mental retardation protein in growth cones via the ubiquitin-proteasome pathway[J]. Front Neural Circuits, 2020, 14: 5.
JIN E J, KO H R, HWANG I, et al. Akt regulates neurite growth by phosphorylation-dependent inhibition of radixin proteasomal degradation[J]. Sci Rep, 2018, 8(1): 2557.
HE L H, LEMASTERS J J. Regulated and unregulated mitochondrial permeability transition pores: a new paradigm of pore structure and function?[J]. FEBS Lett, 2002, 512(1/2/3): 1-7.
MCEWEN M L, SULLIVAN P G, SPRINGER J E. Pretreatment with the cyclosporin derivative, NIM811, improves the function of synaptic mitochondria following spinal cord contusion in rats[J]. J Neurotrauma, 2007, 24(4): 613-624.
PATEL S P, SULLIVAN P G, LYTTLE T S, et al. Acetyl-L-carnitine treatment following spinal cord injury improves mitochondrial function correlated with remarkable tissue sparing and functional recovery[J]. Neuroscience, 2012, 210: 296-307.
WANG Q Q, CAI H X, HU Z X, et al. Loureirin B promotes axon regeneration by inhibiting endoplasmic reticulum stress: induced mitochondrial dysfunction and regulating the Akt/GSK-3β pathway after spinal cord injury[J]. J Neurotrauma, 2019, 36(12): 1949-1964.
SCHOLPA N E, WILLIAMS H, WANG W X, et al. Pharmacological stimulation of mitochondrial biogenesis using the food and drug administration-approved β2-adrenoreceptor agonist formoterol for the treatment of spinal cord injury[J]. J Neurotrauma, 2019, 36(6): 962-972.
GOLLIHUE J L, PATEL S P, RABCHEVSKY A G. Mitochondrial transplantation strategies as potential therapeutics for central nervous system trauma[J]. Neural Regen Res, 2018, 13(2): 194-197.
GOLLIHUE J L, PATEL S P, ELDAHAN K C, et al. Effects of mitochondrial transplantation on bioenergetics, cellular incorporation, and functional recovery after spinal cord injury[J]. J Neurotrauma, 2018, 35(15): 1800-1818.
WU Y Q, XIONG J, HE Z L, et al. Metformin promotes microglial cells to facilitate myelin debris clearance and accelerate nerve repairment after spinal cord injury[J]. Acta Pharmacol Sin, 2022, 43(6): 1360-1371.
BISICCHIA E, MASTRANTONIO R, NOBILI A, et al. Restoration of ER proteostasis attenuates remote apoptotic cell death after spinal cord injury by reducing autophagosome overload[J]. Cell Death Dis, 2022, 13(4): 381.
ZHOU Z J, HU B W, LYU Q N, et al. MiR-384-5p promotes spinal cord injury recovery in rats through suppressing of autophagy and endoplasmic reticulum stress[J]. Neurosci Lett, 2020, 727: 134937.
HE M, DING Y T, CHU C, et al. Autophagy induction stabilizes microtubules and promotes axon regeneration after spinal cord injury[J]. Proc Natl Acad Sci USA, 2016, 113(40): 11324-11329.
LI Y, ZHANG J, ZHOU K L, et al. Elevating sestrin2 attenuates endoplasmic reticulum stress and improves functional recovery through autophagy activation after spinal cord injury[J]. Cell Biol Toxicol, 2021, 37(3): 401-419.
ZHAO Y Z, JIANG X, XIAO J, et al. Using NGF heparin-poloxamer thermosensitive hydrogels to enhance the nerve regeneration for spinal cord injury[J]. Acta Biomater, 2016, 29: 71-80.
LI H T, ZHANG X R, QI X, et al. Icariin inhibits endoplasmic reticulum stress-induced neuronal apoptosis after spinal cord injury through modulating the PI3K/AKT signaling pathway[J]. Int J Biol Sci, 2019, 15(2): 277-286.
LIU X H, YANG J, LI Z, et al. Hyperbaric oxygen treatment protects against spinal cord injury by inhibiting endoplasmic reticulum stress in rats[J]. Spine, 2015, 40(24): E1276-E1283.
WU Q, ZHANG Y J, GAO J Y, et al. Aquaporin-4 mitigates retrograde degeneration of rubrospinal neurons by facilitating edema clearance and glial scar formation after spinal cord injury in mice[J]. Mol Neurobiol, 2014, 49(3): 1327-1337.
ZHENG B B, ZHOU Y L, ZHANG H Y, et al. Dl-3-n-butylphthalide prevents the disruption of blood-spinal cord barrier via inhibiting endoplasmic reticulum stress following spinal cord injury[J]. Int J Biol Sci, 2017, 13(12): 1520-1531.
WANG H L, WU Y Q, HAN W, et al. Hydrogen sulfide ameliorates blood-spinal cord barrier disruption and improves functional recovery by inhibiting endoplasmic reticulum stress-dependent autophagy[J]. Front Pharmacol, 2018, 9: 858.
KOBAYASHI H, ETOH K, FUKUDA M. Rab35 is translocated from Arf6-positive perinuclear recycling endosomes to neurite tips during neurite outgrowth[J]. Small GTPases, 2014, 5(3): e983874.
WU C S, CUI Z M, LIU Y H, et al. The importance of EHD1 in neurite outgrowth contributing to the functional recovery after spinal cord injury[J]. Int J Dev Neurosci, 2016, 52: 24-32.
GALLON M, CLAIRFEUILLE T, STEINBERG F, et al. A unique PDZ domain and arrestin-like fold interaction reveals mechanistic details of endocytic recycling by SNX27-retromer[J]. Proc Natl Acad Sci USA, 2014, 111(35): E3604-E3613.
ZENG Y Z, WANG N W, GUO T T, et al. Snx27 deletion promotes recovery from spinal cord injury by neuroprotection and reduces macrophage/microglia proliferation[J]. Front Neurol, 2018, 9: 1059.
SPITZBARTH I, MOORE S A, STEIN V M, et al. Current insights into the pathology of canine intervertebral disc extrusion-induced spinal cord injury[J]. Front Vet Sci, 2020, 7: 595796.
MYEKU N, WANG H, FIGUEIREDO-PEREIRA M E. cAMP stimulates the ubiquitin/proteasome pathway in rat spinal cord neurons[J]. Neurosci Lett, 2012, 527(2): 126-131.
WU C S, CHEN J J, LIU Y H, et al. Upregulation of PSMB4 is associated with the necroptosis after spinal cord injury[J]. Neurochem Res, 2016, 41(11): 3103-3112.
TICA J R, BRADBURY E J, DIDANGELOS A. Combined transcriptomics, proteomics and bioinformatics identify drug targets in spinal cord injury[J]. Int J Mol Sci, 2018, 19(5): 1461.
... 靶向胞内体治疗能促进脊髓损伤中胞内神经营养物质的运输.Eps15同源结构域蛋白1(Eps15 homology domain protein 1,EHD1)在胞内体系统中发挥着“看门人”的作用[50].在WU等[51]的研究中,EHD1通过循环胞内体而显示出其对神经营养素内吞和运输的调控作用,胞内体跨膜运输也对神经突生长有着促进作用.酪氨酸激酶受体A(tyrosine kinase receptor A,TrkA),作为细胞膜上NGF的功能特异性受体,在与NGF结合时被激活.而EHD1可上调神经元TrkA的表达.该研究[51]显示通过靶向胞内体EHD1,调控TrkA的回收进而提高NGF利用效率,可达到加强对损伤神经的营养物质输送而治疗脊髓损伤的目的.分选连接蛋白27(sorting nexin family member 27,SNX27)是一种胞内体相关的介导分子,参与多种神经疾病的病理和发展过程[52],但其在脊髓损伤中的作用尚待研究.ZENG等[53]的研究发现SNX27在脊髓损伤中上调.该研究者进一步的研究发现SNX27的缺乏对脊髓损伤的神经功能学水平的恢复具有促进作用,且SNX27的缺乏可抑制神经元细胞凋亡.此外,在脊髓损伤的小鼠模型中,SNX27缺乏的小鼠神经损伤部位巨噬细胞/小胶质细胞的增殖受到抑制,并且巨噬细胞/小胶质细胞浸润和活化降低.该研究[53]提示下调SNX27是一种靶向急性神经元死亡和慢性神经炎症的潜在疗法,并有助于促进脊髓损伤后的神经组织学和功能学水平的恢复,为靶向胞内体治疗脊髓损伤的潜在靶点. ...
2
... 靶向胞内体治疗能促进脊髓损伤中胞内神经营养物质的运输.Eps15同源结构域蛋白1(Eps15 homology domain protein 1,EHD1)在胞内体系统中发挥着“看门人”的作用[50].在WU等[51]的研究中,EHD1通过循环胞内体而显示出其对神经营养素内吞和运输的调控作用,胞内体跨膜运输也对神经突生长有着促进作用.酪氨酸激酶受体A(tyrosine kinase receptor A,TrkA),作为细胞膜上NGF的功能特异性受体,在与NGF结合时被激活.而EHD1可上调神经元TrkA的表达.该研究[51]显示通过靶向胞内体EHD1,调控TrkA的回收进而提高NGF利用效率,可达到加强对损伤神经的营养物质输送而治疗脊髓损伤的目的.分选连接蛋白27(sorting nexin family member 27,SNX27)是一种胞内体相关的介导分子,参与多种神经疾病的病理和发展过程[52],但其在脊髓损伤中的作用尚待研究.ZENG等[53]的研究发现SNX27在脊髓损伤中上调.该研究者进一步的研究发现SNX27的缺乏对脊髓损伤的神经功能学水平的恢复具有促进作用,且SNX27的缺乏可抑制神经元细胞凋亡.此外,在脊髓损伤的小鼠模型中,SNX27缺乏的小鼠神经损伤部位巨噬细胞/小胶质细胞的增殖受到抑制,并且巨噬细胞/小胶质细胞浸润和活化降低.该研究[53]提示下调SNX27是一种靶向急性神经元死亡和慢性神经炎症的潜在疗法,并有助于促进脊髓损伤后的神经组织学和功能学水平的恢复,为靶向胞内体治疗脊髓损伤的潜在靶点. ...
... [51]显示通过靶向胞内体EHD1,调控TrkA的回收进而提高NGF利用效率,可达到加强对损伤神经的营养物质输送而治疗脊髓损伤的目的.分选连接蛋白27(sorting nexin family member 27,SNX27)是一种胞内体相关的介导分子,参与多种神经疾病的病理和发展过程[52],但其在脊髓损伤中的作用尚待研究.ZENG等[53]的研究发现SNX27在脊髓损伤中上调.该研究者进一步的研究发现SNX27的缺乏对脊髓损伤的神经功能学水平的恢复具有促进作用,且SNX27的缺乏可抑制神经元细胞凋亡.此外,在脊髓损伤的小鼠模型中,SNX27缺乏的小鼠神经损伤部位巨噬细胞/小胶质细胞的增殖受到抑制,并且巨噬细胞/小胶质细胞浸润和活化降低.该研究[53]提示下调SNX27是一种靶向急性神经元死亡和慢性神经炎症的潜在疗法,并有助于促进脊髓损伤后的神经组织学和功能学水平的恢复,为靶向胞内体治疗脊髓损伤的潜在靶点. ...
1
... 靶向胞内体治疗能促进脊髓损伤中胞内神经营养物质的运输.Eps15同源结构域蛋白1(Eps15 homology domain protein 1,EHD1)在胞内体系统中发挥着“看门人”的作用[50].在WU等[51]的研究中,EHD1通过循环胞内体而显示出其对神经营养素内吞和运输的调控作用,胞内体跨膜运输也对神经突生长有着促进作用.酪氨酸激酶受体A(tyrosine kinase receptor A,TrkA),作为细胞膜上NGF的功能特异性受体,在与NGF结合时被激活.而EHD1可上调神经元TrkA的表达.该研究[51]显示通过靶向胞内体EHD1,调控TrkA的回收进而提高NGF利用效率,可达到加强对损伤神经的营养物质输送而治疗脊髓损伤的目的.分选连接蛋白27(sorting nexin family member 27,SNX27)是一种胞内体相关的介导分子,参与多种神经疾病的病理和发展过程[52],但其在脊髓损伤中的作用尚待研究.ZENG等[53]的研究发现SNX27在脊髓损伤中上调.该研究者进一步的研究发现SNX27的缺乏对脊髓损伤的神经功能学水平的恢复具有促进作用,且SNX27的缺乏可抑制神经元细胞凋亡.此外,在脊髓损伤的小鼠模型中,SNX27缺乏的小鼠神经损伤部位巨噬细胞/小胶质细胞的增殖受到抑制,并且巨噬细胞/小胶质细胞浸润和活化降低.该研究[53]提示下调SNX27是一种靶向急性神经元死亡和慢性神经炎症的潜在疗法,并有助于促进脊髓损伤后的神经组织学和功能学水平的恢复,为靶向胞内体治疗脊髓损伤的潜在靶点. ...
2
... 靶向胞内体治疗能促进脊髓损伤中胞内神经营养物质的运输.Eps15同源结构域蛋白1(Eps15 homology domain protein 1,EHD1)在胞内体系统中发挥着“看门人”的作用[50].在WU等[51]的研究中,EHD1通过循环胞内体而显示出其对神经营养素内吞和运输的调控作用,胞内体跨膜运输也对神经突生长有着促进作用.酪氨酸激酶受体A(tyrosine kinase receptor A,TrkA),作为细胞膜上NGF的功能特异性受体,在与NGF结合时被激活.而EHD1可上调神经元TrkA的表达.该研究[51]显示通过靶向胞内体EHD1,调控TrkA的回收进而提高NGF利用效率,可达到加强对损伤神经的营养物质输送而治疗脊髓损伤的目的.分选连接蛋白27(sorting nexin family member 27,SNX27)是一种胞内体相关的介导分子,参与多种神经疾病的病理和发展过程[52],但其在脊髓损伤中的作用尚待研究.ZENG等[53]的研究发现SNX27在脊髓损伤中上调.该研究者进一步的研究发现SNX27的缺乏对脊髓损伤的神经功能学水平的恢复具有促进作用,且SNX27的缺乏可抑制神经元细胞凋亡.此外,在脊髓损伤的小鼠模型中,SNX27缺乏的小鼠神经损伤部位巨噬细胞/小胶质细胞的增殖受到抑制,并且巨噬细胞/小胶质细胞浸润和活化降低.该研究[53]提示下调SNX27是一种靶向急性神经元死亡和慢性神经炎症的潜在疗法,并有助于促进脊髓损伤后的神经组织学和功能学水平的恢复,为靶向胞内体治疗脊髓损伤的潜在靶点. ...



{kind=link}
{kind=link}