Bioinformatics analysis of pathological mechanism of degenerated tendon via stress deprivation
LI Qinglin,1, WANG Wenbo,2, LIU Wei,2
1.Plastic Surgery Research Institute, Weifang Medical University, Weifang 261042, China
2.Department of Plastic and Reconstructive Surgery, Shanghai Ninth People′s Hospital, Shanghai Jiao Tong University School of Medicine; Shanghai Tissue Engineering Key Laboratory, Shanghai 200011, China
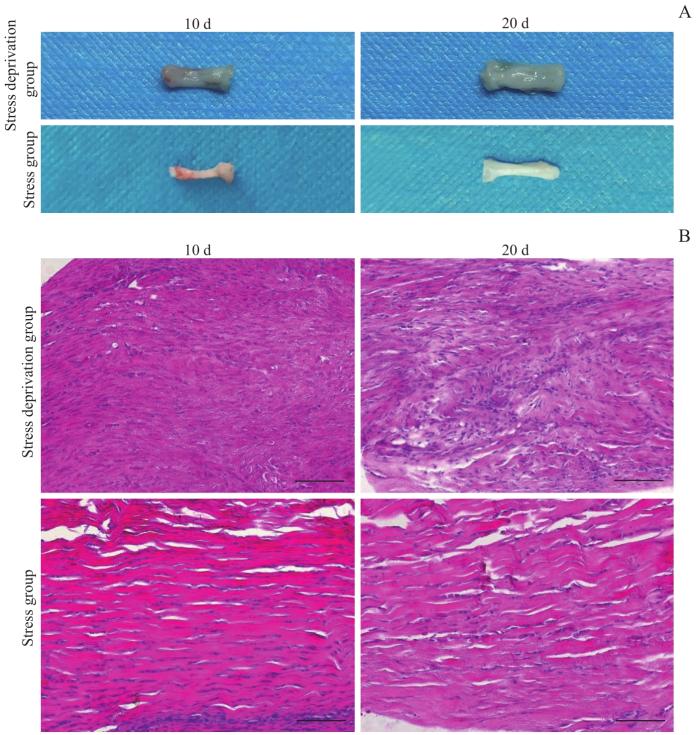
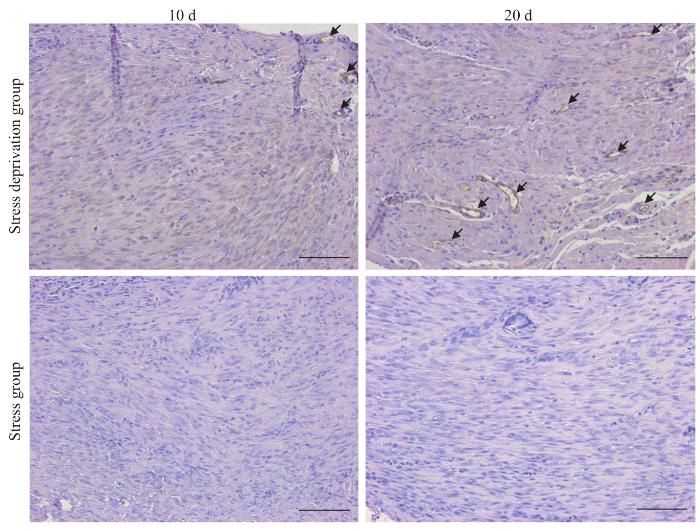
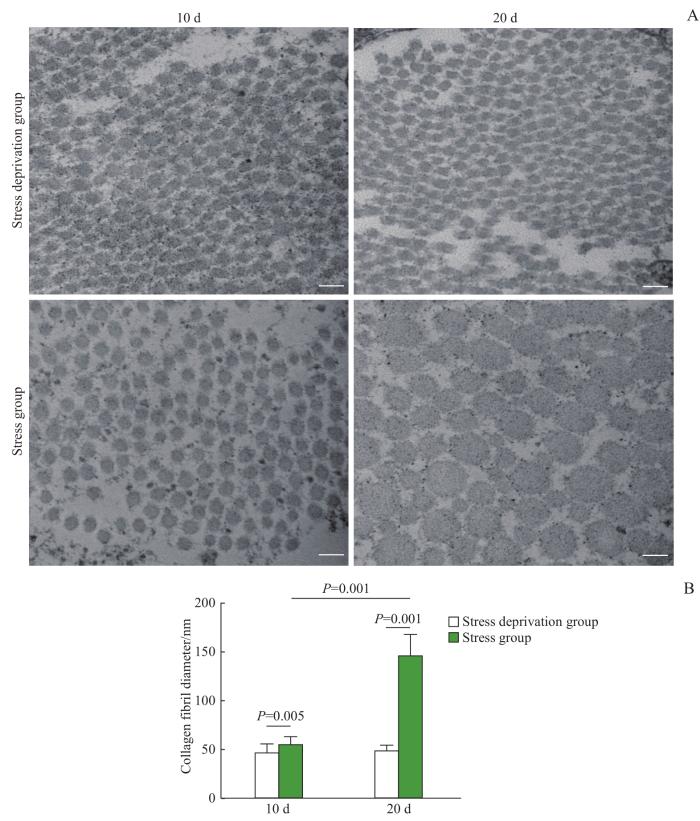
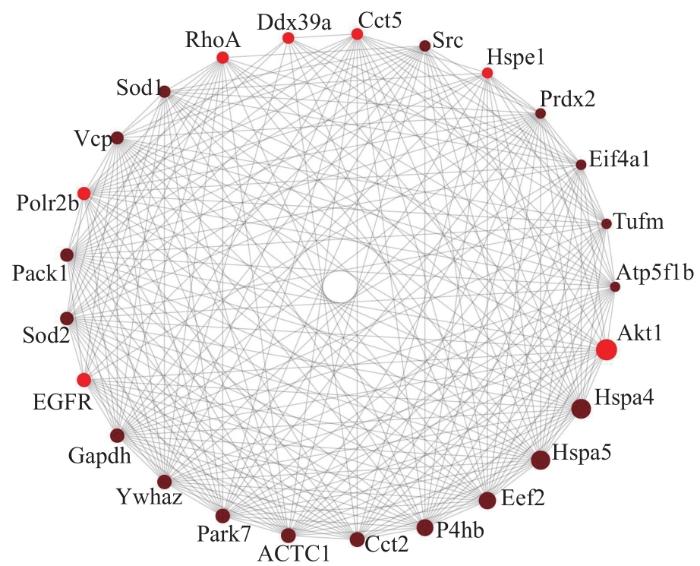
Objective ·To explore relevant molecular mechanisms of the stress deprivation model of newborn rats by using bioinformatics analysis. Methods ·A total of 60 SD rats (10 d post-natal) were enrolled in the study. Those the left Achilles tendon of which was severed were chosen as the experimental group (stress deprivation group), and the right Achilles tendon of which was injured by clamping were chosen as the control group (stress group). On the 10th and 20th day after treatment, tissue samples were collected for gross observation of the tendon development, histological staining of the tendon structure, transmission electron microscope observation of the tendon ultrastructure and immunohistochemical analysis of CD31 expression. Differentially expressed proteins between the two groups at two time points were obtained by using protein mass spectrometry, and GO and KEGG enrichment analysis as well as protein-protein interaction (PPI) network anlysis were performed on differential proteins. Results ·The stress-deprived tendon showed tissue enlargement and congestion, disorganized tendon tissue structure and immature collagen fibers. Transmission electron microscopy showed that the development and maturation of collagen fibrils were significantly impaired in the experimental group, and the diameter of collagen fibrils of the severed tendon became thinner on the 20th day after treatment (P=0.001). Immunohistochemistry showed that the severed tendon was relatively vascularized. The results of protein mass spectrometry analysis showed there were 1 865 and 965 differentially expressed proteins on the 10th and 20th day after treatment, including 1 835 and 837 upregulated proteins respectively. GO analysis showed that the upregulated proteins were involved in biological processes such as intracellular protein transport, protein stabilization, mRNA splicing via spliceosome, protein folding and protein import into nucleus. KEGG analysis indentified enhancement of vascular endothelial growth factor (VEGF) signal pathway, mammalian target of rapamycin (mTOR) signal pathway, endocytosis and other signal pathways in the experimental group. PPI network analysis showed various upregulated proteins including Akt1, Hspa4, Hspa5, Eef2, ACTC1 and RhoA. Conclusion ·Stress deprivation can activate multiple signal pathways in tendon cells and lead to tissue vascularization, abnormal collagen development, etc., resulting in degenerative pathological changes of tendons.
LI Qinglin, WANG Wenbo, LIU Wei. Bioinformatics analysis of pathological mechanism of degenerated tendon via stress deprivation. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(5): 560-570 doi:10.3969/j.issn.1674-8115.2023.05.006
使用R Studio 1.1.463软件进行统计分析和绘图。将LC-MS/MS原始文件导入Spectronaut Pulsar软件进行搜库建库,使用Spectronaut Pulsar软件的“Analysis”模块,选择“+”新建分析;根据软件提示逐步完成参数设置,开始分析;分析完成后,进入“Report”模块导出定量数据。将表达值≥50%的蛋白样本保留,用同一组样品的均值填充缺失值≤50%的蛋白。通过log2转换和中位数归一化获得可信蛋白。基于获得的可信蛋白,选择2种不同标准计算样本之间的差值。在处理数据时,用1.5倍数变化评估样本之间某一蛋白表达水平倍数变化。
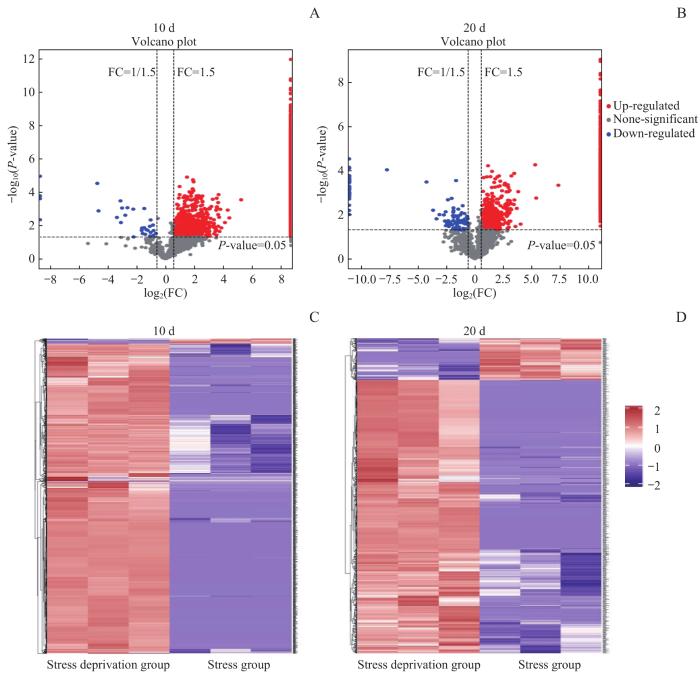
Note: A/B. Differentially expressed protein volcano maps on the 10th (A) and 20th (B) day. C/D. Differential expression cluster heat maps on the 10th (C) and 20th (D) day.
Fig 4
Volcanic diagram and cluster thermogram of differentially expressed proteins between tendons with and without mechanical loading
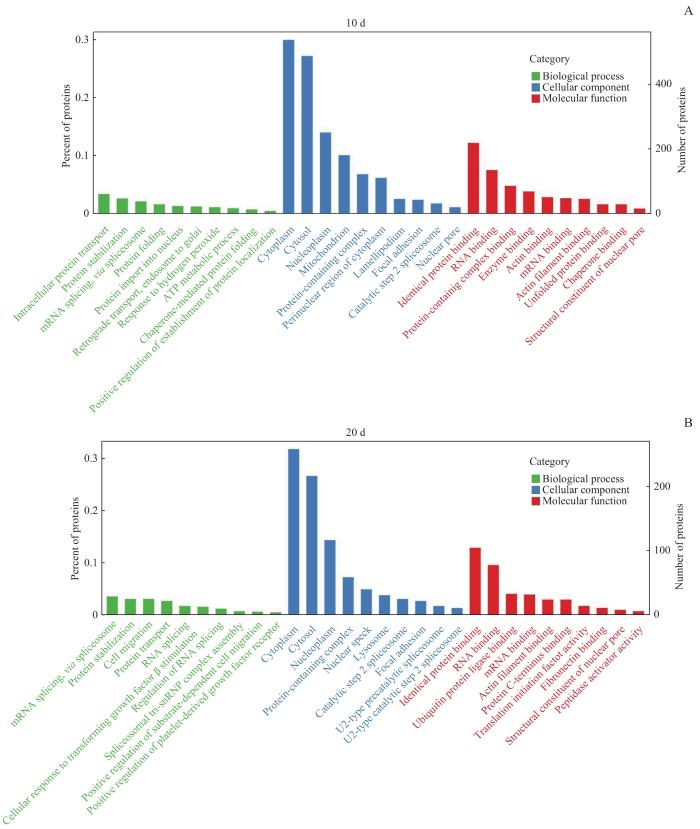
Note: A and B represent the differentially expressed protein GO analysis between stress deprivation and stress-stimulated tendons on the 10th and 20th day after surgery, respectively.
Fig 5
GO analysis of differentially expressed proteins between tendons with and without mechanical loading
2.6 KEGG分析
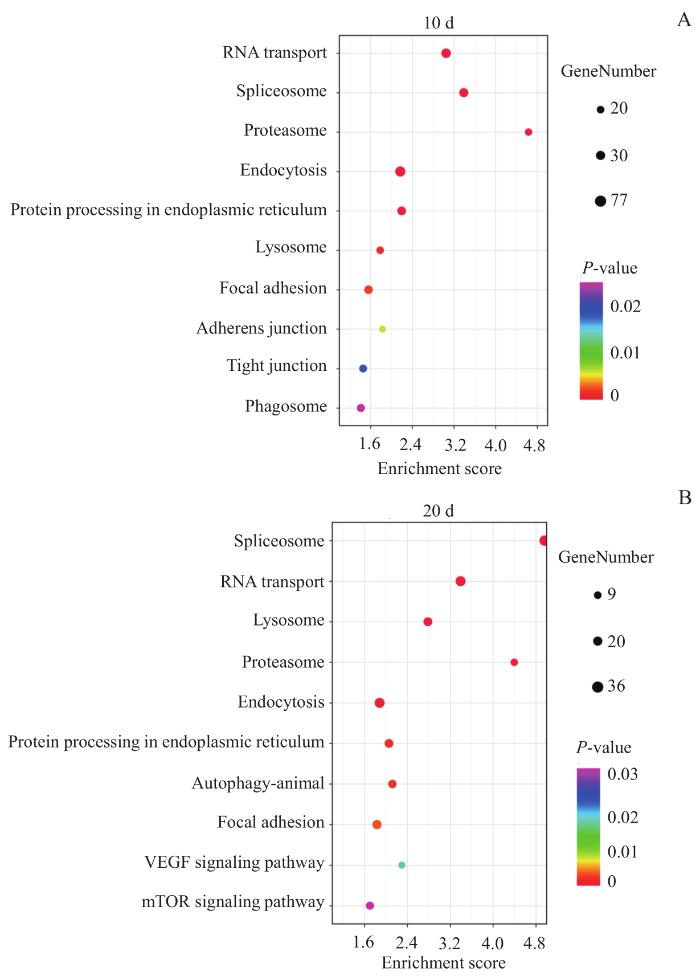
将手术处理后第10日和第20日上调差异蛋白分别进行KEGG通路富集,结果显示:处理后第10日富集到310条信号通路,处理后第20日富集到289条信号通路。对2次富集到的信号通路进行筛选(图6),发现排名靠前的与肌腱胶原退行病理改变相关通路包括溶酶体、黏着斑、自噬、血管内皮生长因子(vascular endothelial growth factor,VEGF)信号通路、哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)信号通路、吞噬作用等通路。
Note: A and B represent KEGG analysis of differentially expressed protein between stress deprivation and stress-stimulated tendons on the 10th and 20th day after surgery, respectively.
Fig 6
KEGG analysis of differentially expressed proteins between tendon with and without mechanical loading
LI Qinglin, LIU Wei and WANG Wenbo were responsible for conceptualization. Experimental design and verification were done by LIU Wei. Data analysis and collation were done by LI Qinglin and WANG Wenbo. Original draft was written by LI Qinglin. Manuscript review and editing were done by LIU Wei and WANG Wenbo. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
HEO S J, SZCZESNY S E, KIM D H, et al. Expansion of mesenchymal stem cells on electrospun scaffolds maintains stemness, mechano-responsivity, and differentiation potential[J]. J Orthop Res, 2018, 36(2): 808-815.
FRIZZIERO A, FINI M, SALAMANNA F, et al. Effect of training and sudden detraining on the patellar tendon and its enthesis in rats[J]. BMC Musculoskelet Disord, 2011, 12: 20.
WUNDERLI S L, BLACHE U, BERETTA PICCOLI A, et al. Tendon response to matrix unloading is determined by the patho-physiological niche[J]. Matrix Biol, 2020, 89: 11-26.
CHEN Z, ZHOU B, WANG X, et al. Synergistic effects of mechanical stimulation and crimped topography to stimulate natural collagen development for tendon engineering[J]. Acta Biomater, 2022, 145: 297-315.
SMYTH L M, TAMURA K, OLIVEIRA M, et al. Capivasertib, an AKT kinase inhibitor, as mnotherapy or in combination with fulvestrant in patients with AKT1 E17K-mutant, ER-positive metastatic breast cancer[J]. Clin Cancer Res, 2020, 26(15): 3947-3957.
FUJIMOTO Y, MORITA T Y, OHASHI A, et al. Combination treatment with a PI3K/Akt/mTOR pathway inhibitor overcomes resistance to anti-HER2 therapy in PIK3CA-mutant HER2-positive breast cancer cells[J]. Sci Rep, 2020, 10(1): 21762.
CONG X X, RAO X S, LIN J X, et al. Activation of AKT-mTOR signaling directs tenogenesis of mesenchymal stem cells[J]. Stem Cells, 2018, 36(4): 527-539.
ZASECK L W, MILLER R A, BROOKS S V. Rapamycin attenuates age-associated changes in tibialis anterior tendon viscoelastic properties[J]. J Gerontol A Biol Sci Med Sci, 2016, 71(7): 858-865.
TUMMINIA S J, MITTON K P, ARORA J, et al. Mechanical stretch alters the actin cytoskeletal network and signal transduction in human trabecular meshwork cells[J]. Invest Ophthalmol Vis Sci, 1998, 39(8): 1361-1371.
SARASA-RENEDO A, TUNÇ-CIVELEK V, CHIQUET M. Role of RhoA/ROCK-dependent actin contractility in the induction of tenascin-C by cyclic tensile strain[J]. Exp Cell Res, 2006, 312(8): 1361-1370.
RIDLEY A J, HALL A. The small GTP-binding protein rho regulates the assembly of focal adhesions and actin stress fibers in response to growth factors[J]. Cell, 1992, 70(3): 389-399.
TERAMURA T, TAKEHARA T, ONODERA Y, et al. Mechanical stimulation of cyclic tensile strain induces reduction of pluripotent related gene expressions via activation of Rho/ROCK and subsequent decreasing of AKT phosphorylation in human induced pluripotent stem cells[J]. Biochem Biophys Res Commun, 2012, 417(2): 836-841.
TSAO J, VERNET D A, GELFAND R, et al. Myostatin genetic inactivation inhibits myogenesis by muscle-derived stem cells in vitro but not when implanted in the mdx mouse muscle[J]. Stem Cell Res Ther, 2013, 4(1): 4.
LI A, SU X, TIAN Y, et al. Effect of actin alpha cardiac muscle 1 on the proliferation and differentiation of bovine myoblasts and preadipocytes[J]. Animals (Basel), 2021, 11(12): 3468.
PUFE T, PETERSEN W J, MENTLEIN R, et al. The role of vasculature and angiogenesis for the pathogenesis of degenerative tendons disease[J]. Scand J Med Sci Sports, 2005, 15(4): 211-222.
PERRY S M, MCILHENNY S E, HOFFMAN M C, et al. Inflammatory and angiogenic mRNA levels are altered in a supraspinatus tendon overuse animal model[J]. J Shoulder Elbow Surg, 2005, 14(1 suppl s): 79S-83S.
HU J, DING X, TIAN S, et al. TRIM39 deficiency inhibits tumor progression and autophagic flux in colorectal cancer via suppressing the activity of Rab7[J]. Cell Death Dis, 2021, 12(4): 391.
XU K, LIN C, MA D, et al. Spironolactone ameliorates senescence and calcification by modulating autophagy in rat tendon-derived stem cells via the NF-κB/MAPK pathway[J]. Oxid Med Cell Longev, 2021, 2021: 5519587.
WANG H, GAO N, LI Z, et al. Autophagy alleviates melamine-induced cell death in PC12 cells via decreasing ROS level[J]. Mol Neurobiol, 2016, 53(3): 1718-1729.
MARIÑO G, NISO-SANTANO M, BAEHRECKE E H, et al. Self-consumption: the interplay of autophagy and apoptosis[J]. Nat Rev Mol Cell Biol, 2014, 15(2): 81-94.
ARBI S, EKSTEEN E C, OBERHOLZER H M, et al. Premature collagen fibril formation, fibroblast-mast cell interactions and mast cell-mediated phagocytosis of collagen in keloids[J]. Ultrastruct Pathol, 2015, 39(2): 95-103.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}