Osteoporosis (OP) is a chronic senile bone disease characterized by decreased bone mass and increased bone fragility. There are many inducing factors and the pathogenesis is complex. To explore the mechanism of OP and improve clinical efficacy has always been a hot topic in life science. In recent years, it has been found that mitochondria play an important role in the pathogenesis of OP. Functional abnormalities such as mitochondrial energy metabolism, mitochondrial oxidative stress, mitochondrial autophagy, mitochondrial-mediated apoptosis and mitochondrial dynamics can interfere with the differentiation of bone marrow mesenchymal stem cells through different signal pathways, cytokines and protein expression to regulate osteoblast activity, proliferation and differentiation, and start the process of osteoclast apoptosis. Therefore, taking mitochondria as the target, regulating the functions of mitochondrial energy metabolism, oxidative stress, autophagy and kinetics, inducing osteogenic differentiation of bone marrow mesenchymal stem cells, promoting osteoblast differentiation and mineralization, and inducing osteoclast apoptosis are potential strategies for the prevention and treatment of OP. In this article, the mechanism of mitochondrial dysfunction in OP was reviewed by referring to relevant literature at home and abroad, in order to lay a foundation for further research.
JIN Fangquan, FAN Chenghu, TANG Xiaodong, CHEN Yantong, QI Bingxian. Research progress in the relationship between mitochondrial dysfunction and osteoporosis. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(6): 761-767 doi:10.3969/j.issn.1674-8115.2023.06.013
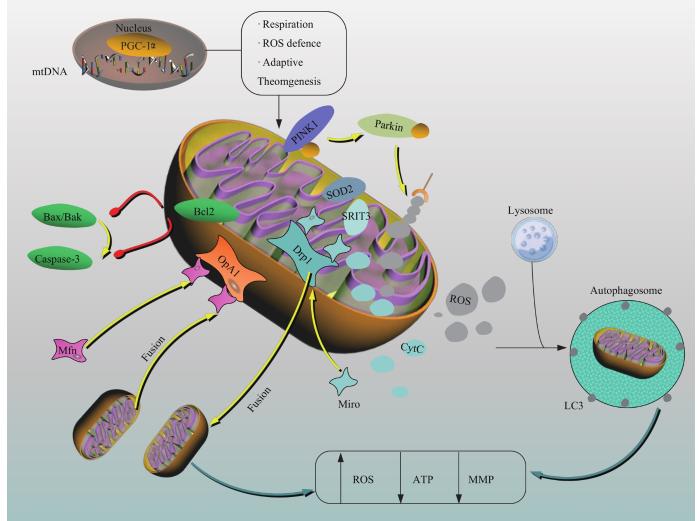
Fig1
Mechanism and key factors of mitochondrial dysfunction in OP
2.1 线粒体能量代谢与OP
细胞中产生ATP的方式主要有3种:一是糖酵解,二是TCA,三是线粒体OXPHOS。糖酵解和TCA产生还原型烟酰胺腺嘌呤二核苷酸、还原型黄素腺嘌呤二核苷酸和其他分子,而OXPHOS利用这些物质还原氧气并合成ATP[16]。BMSCs增殖和自我更新时通过糖酵解获取能量,在分化时通过OXPHOS和TCA产生更多ATP,这可能是因为无氧代谢可避免ROS对mtDNA和细胞膜造成氧化损伤[17]。在BMSCs成骨分化时线粒体膜蛋白和mtDNA数量显著增加,PGC-1α表达上调,促进OXPHOS产生ATP为其供能[18]。抑制糖酵解时,线粒体呼吸减弱,ROS产生减少,而细胞内ROS减少可降低氧化应激损伤,激活神经源性位点缺口同源蛋白(neurogenic loci gap homologous protein,Notch)信号通路,增加BMSCs成骨分化,促进骨形成[19]。HIF-1α是调节线粒体代谢途径的关键转录因子,在TCA中下调HIF-1α会抑制骨细胞对葡萄糖的摄取和OC对骨吸收[20]。因此,调节mtDNA数量和HIF-1α、PGC-1α基因表达,改变线粒体能量代谢途径,诱导BMSCs成骨分化可治疗OP。
细胞凋亡是调节细胞死亡的一种主要形式。线粒体介导的凋亡是指B细胞淋巴瘤2(B cell lymphoma 2,Bcl-2)调控关闭Bcl-2相关X蛋白(Bcl-2 associated X protein,Bax)在线粒体外膜上形成的通道,阻断细胞色素C释放,降低线粒体外膜通透性,激活半胱氨酸天冬氨酸蛋白酶(cysteinyl aspartate specific proteinase,caspase)3和7,调控细胞凋亡[35]。随着生物科技发展,BMSCs移植已广泛应用于组织修复和再生,但因移植过程中干细胞的高凋亡率而使其保存技术亟需改进[36]。Bcl-2和Bax的比率决定了线粒体介导的细胞凋亡后果[37],近期有研究佐证这一观点[38]。含镁基质中BMSCs内SOD2和CAT活性增强,JC-1染色后观察到MMP升高,PCR检测Bcl-2/Bax的比率升高,BMSCs凋亡被抑制。因此,提高Bcl-2/Bax比率,抑制骨形成相关细胞凋亡是防治OP的新思路。
线粒体动力学是指线粒体动态分裂和融合。通过分裂增加其数量,维持细胞极性,消除受损的线粒体,该过程由线粒体动力相关蛋白1(dynamin-related protein 1,Drp1)和线粒体分裂因子调控;融合时线粒体进行内容物交换和连接,提供ATP,减轻氧化损伤,并维持MMP,该过程由线粒体外膜的视神经萎缩蛋白1(optic atrophy protein-1,OPA1)、内膜丝裂蛋白介导[41]。线粒体动力学通过协调能量供应、细胞内ROS产生和钙平衡来调节细胞命运。在BMSCs分化、成熟的过程中,启动OXPHOS,线粒体延长并且互相连接,Drp1表达增加,OPA1表达下降,在聚丙烯酸与胶原膜复合体基质中,矿化结节增多,这说明线粒体分裂有利于BMSCs向成骨分化并矿化[42-43]。与之相反,BMSCs凋亡伴随线粒体异常分裂,过度分裂发生时,细胞内Ca2+内流,导致β-catenin蛋白降解,损害BMSCs分化功能[44]。
JIN Fangquan put forward the innovation of the article, collected and sorted out the literature, and was responsible for writing. TANG Xiaodong and FAN Chenghu conducted the article guidance, revision and financial support. CHEN Yantong participated in the literature inquiry. QI Bingxian participated in the revision of the paper and the adjustment of the article frame structure. All the authors read and agreed to the submission of the final manuscript.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
JIANG Y H, ZHANG P, ZHANG X, et al. Advances in mesenchymal stem cell transplantation for the treatment of osteoporosis[J]. Cell Prolif, 2021, 54(1): e12956.
WU H Y, LIU Y, LAN Y J, et al. Meta-analysis of the prevalence of osteoporosis in Chinese postmenopausal women[J]. Chinese Journal of Evidence-Based Medicine, 2022, 22(8):882-890.
CUI L J, JACKSON M, WESSLER Z, et al. Estimating the future clinical and economic benefits of improving osteoporosis diagnosis and treatment among women in China: a simulation projection model from 2020 to 2040[J]. Arch Osteoporos, 2021, 16(1): 118.
YAN W H, DIAO S, FAN Z P. The role and mechanism of mitochondrial functions and energy metabolism in the function regulation of the mesenchymal stem cells[J]. Stem Cell Res Ther, 2021, 12(1): 140.
DONAT A, KNAPSTEIN P R, JIANG S, et al. Glucose metabolism in osteoblasts in healthy and pathophysiological conditions[J]. Int J Mol Sci, 2021, 22(8): 4120.
LI T C, YAN Z Q, HE S S, et al. Intermittent parathyroid hormone improves orthodontic retention via insulin-like growth factor-1[J]. Oral Dis, 2021, 27(2): 290-300.
ANGIREDDY R, KAZMI H R, SRINIVASAN S, et al. Cytochrome c oxidase dysfunction enhances phagocytic function and osteoclast formation in macrophages[J]. FASEB J, 2019, 33(8): 9167-9181.
ZHANG Y, ROHATGI N, VEIS D J, et al. PGC1β organizes the osteoclast cytoskeleton by mitochondrial biogenesis and activation[J]. J Bone Miner Res, 2018, 33(6): 1114-1125.
GUO L, CHEN K Z, YUAN J, et al. Estrogen inhibits osteoclasts formation and bone resorption via microRNA-27a targeting PPARγ and APC[J]. J Cell Physiol, 2018, 234(1): 581-594.
GUO Y S, CHI X P, WANG Y F, et al. Mitochondria transfer enhances proliferation, migration, and osteogenic differentiation of bone marrow mesenchymal stem cell and promotes bone defect healing[J]. Stem Cell Res Ther, 2020, 11(1): 245.
YAN W H, DIAO S, FAN Z P. The role and mechanism of mitochondrial functions and energy metabolism in the function regulation of the mesenchymal stem cells[J]. Stem Cell Res Ther, 2021, 12(1): 140.
FENG X R, ZHANG W J, YIN W, et al. The involvement of mitochondrial fission in maintenance of the stemness of bone marrow mesenchymal stem cells[J]. Exp Biol Med (Maywood), 2019, 244(1): 64-72.
JOHNSON J, MERCADO-AYON E, MERCADO-AYON Y, et al. Mitochondrial dysfunction in the development and progression of neurodegenerative diseases[J]. Arch Biochem Biophys, 2021, 702: 108698.
JIN Y, SHEN Y, SU X, et al. The small GTPases Rab27b regulates mitochondrial fatty acid oxidative metabolism of cardiac mesenchymal stem cells[J]. Front Cell Dev Biol, 2020, 8: 209.
PAL S, SINGH M, PORWAL K, et al. Adiponectin receptors by increasing mitochondrial biogenesis and respiration promote osteoblast differentiation: discovery of isovitexin as a new class of small molecule adiponectin receptor modulator with potential osteoanabolic function[J]. Eur J Pharmacol, 2021, 913: 174634.
LEE S Y, LONG F X. Notch signaling suppresses glucose metabolism in mesenchymal progenitors to restrict osteoblast differentiation[J]. J Clin Invest, 2018, 128(12): 5573-5586.
KNOWLES H J. Distinct roles for the hypoxia-inducible transcription factors HIF-1α and HIF-2α in human osteoclast formation and function[J]. Sci Rep, 2020, 10(1): 21072.
SHARES B H, BUSCH M, WHITE N, et al. Active mitochondria support osteogenic differentiation by stimulating β-catenin acetylation[J]. J Biol Chem, 2018, 293(41): 16019-16027.
KUSHWAHA P, ALEKOS N S, KIM S P, et al. Mitochondrial fatty acid β-oxidation is important for normal osteoclast formation in growing female mice[J]. Front Physiol, 2022, 13: 997358.
LI X M, CHEN Y, MAO Y X, et al. Curcumin protects osteoblasts from oxidative stress-induced dysfunction via GSK3β-Nrf2 signaling pathway[J]. Front Bioeng Biotechnol, 2020, 8: 625.
MA C, SUN Y N, PI C C, et al. Sirt3 attenuates oxidative stress damage and rescues cellular senescence in rat bone marrow mesenchymal stem cells by targeting superoxide dismutase 2[J]. Front Cell Dev Biol, 2020, 8: 599376.
CAO X C, LUO D Q, LI T, et al. MnTBAP inhibits bone loss in ovariectomized rats by reducing mitochondrial oxidative stress in osteoblasts[J]. J Bone Miner Metab, 2020, 38(1): 27-37.
LIU H D, REN M X, LI Y, et al. Melatonin alleviates hydrogen peroxide induced oxidative damage in MC3T3-E1 cells and promotes osteogenesis by activating SIRT1[J]. Free Radic Res, 2022, 56(1): 63-76.
KIM H N, PONTE F, NOOKAEW I, et al. Estrogens decrease osteoclast number by attenuating mitochondria oxidative phosphorylation and ATP production in early osteoclast precursors[J]. Sci Rep, 2020, 10(1): 11933.
FENG X R, YIN W, WANG J L, et al. Mitophagy promotes the stemness of bone marrow-derived mesenchymal stem cells[J]. Exp Biol Med (Maywood), 2021, 246(1): 97-105.
FAN P, YU X Y, XIE X H, et al. Mitophagy is a protective response against oxidative damage in bone marrow mesenchymal stem cells[J]. Life Sci, 2019, 229: 36-45.
GUO Y Y, JIA X, CUI Y Z, et al. Sirt3-mediated mitophagy regulates AGEs-induced BMSCs senescence and senile osteoporosis[J]. Redox Biol, 2021, 41: 101915.
WANG X D, MA H D, SUN J, et al. Mitochondrial ferritin deficiency promotes osteoblastic ferroptosis via mitophagy in type 2 diabetic osteoporosis[J]. Biol Trace Elem Res, 2022, 200(1): 298-307.
AOKI S, SHIMIZU K, ITO K. Autophagy-dependent mitochondrial function regulates osteoclast differentiation and maturation[J]. Biochem Biophys Res Commun, 2020, 527(4): 874-880.
QIU T, HE Y Y, ZHANG X, et al. Novel role of ER stress and mitochondria stress in serum-deprivation induced apoptosis of rat mesenchymal stem cells[J]. Curr Med Sci, 2018, 38(2): 229-235.
CHEN Y M, XIONG S B, ZHAO F H, et al. Effect of magnesium on reducing the UV-induced oxidative damage in marrow mesenchymal stem cells[J]. J Biomed Mater Res A, 2019, 107(6): 1253-1263.
YANG K D, PEI L, ZHOU S M, et al. Metformin attenuates H2O2-induced osteoblast apoptosis by regulating SIRT3 via the PI3K/AKT pathway[J]. Exp Ther Med, 2021, 22(5): 1316.
ZHENG D L, CUI C L, SHAO C, et al. Coenzyme Q10 inhibits RANKL-induced osteoclastogenesis by regulation of mitochondrial apoptosis and oxidative stress in RAW264.7 cells[J]. J Biochem Mol Toxicol, 2021, 35(7): e22778.
REN L, CHEN X D, CHEN X B, et al. Mitochondrial dynamics: fission and fusion in fate determination of mesenchymal stem cells[J]. Front Cell Dev Biol, 2020, 8: 580070.
WAN M C, TANG X Y, LI J, et al. Upregulation of mitochondrial dynamics is responsible for osteogenic differentiation of mesenchymal stem cells cultured on self-mineralized collagen membranes[J]. Acta Biomater, 2021, 136: 137-146.
FENG X R, ZHANG W J, YIN W, et al. The involvement of mitochondrial fission in maintenance of the stemness of bone marrow mesenchymal stem cells[J]. Exp Biol Med (Maywood), 2019, 244(1): 64-72.
ZHONG X Y, CUI P, CAI Y P, et al. Mitochondrial dynamics is critical for the full pluripotency and embryonic developmental potential of pluripotent stem cells[J]. Cell Metab, 2019, 29(4): 979-992.e4.
PAHWA H, KHAN M T, SHARAN K. Hyperglycemia impairs osteoblast cell migration and chemotaxis due to a decrease in mitochondrial biogenesis[J]. Mol Cell Biochem, 2020, 469(1/2): 109-118.
JEONG S, SEONG J H, KANG J H, et al. Dynamin-related protein 1 positively regulates osteoclast differentiation and bone loss[J]. FEBS Lett, 2021, 595(1): 58-67.
NISHIKAWA K, TAKEGAMI H, SESAKI H. Opa1-mediated mitochondrial dynamics is important for osteoclast differentiation[J]. MicroPubl Biol, 2022. DOI: 10.17912/micropub.biology.000650.
... 细胞中产生ATP的方式主要有3种:一是糖酵解,二是TCA,三是线粒体OXPHOS.糖酵解和TCA产生还原型烟酰胺腺嘌呤二核苷酸、还原型黄素腺嘌呤二核苷酸和其他分子,而OXPHOS利用这些物质还原氧气并合成ATP[16].BMSCs增殖和自我更新时通过糖酵解获取能量,在分化时通过OXPHOS和TCA产生更多ATP,这可能是因为无氧代谢可避免ROS对mtDNA和细胞膜造成氧化损伤[17].在BMSCs成骨分化时线粒体膜蛋白和mtDNA数量显著增加,PGC-1α表达上调,促进OXPHOS产生ATP为其供能[18].抑制糖酵解时,线粒体呼吸减弱,ROS产生减少,而细胞内ROS减少可降低氧化应激损伤,激活神经源性位点缺口同源蛋白(neurogenic loci gap homologous protein,Notch)信号通路,增加BMSCs成骨分化,促进骨形成[19].HIF-1α是调节线粒体代谢途径的关键转录因子,在TCA中下调HIF-1α会抑制骨细胞对葡萄糖的摄取和OC对骨吸收[20].因此,调节mtDNA数量和HIF-1α、PGC-1α基因表达,改变线粒体能量代谢途径,诱导BMSCs成骨分化可治疗OP. ...
1
... 细胞中产生ATP的方式主要有3种:一是糖酵解,二是TCA,三是线粒体OXPHOS.糖酵解和TCA产生还原型烟酰胺腺嘌呤二核苷酸、还原型黄素腺嘌呤二核苷酸和其他分子,而OXPHOS利用这些物质还原氧气并合成ATP[16].BMSCs增殖和自我更新时通过糖酵解获取能量,在分化时通过OXPHOS和TCA产生更多ATP,这可能是因为无氧代谢可避免ROS对mtDNA和细胞膜造成氧化损伤[17].在BMSCs成骨分化时线粒体膜蛋白和mtDNA数量显著增加,PGC-1α表达上调,促进OXPHOS产生ATP为其供能[18].抑制糖酵解时,线粒体呼吸减弱,ROS产生减少,而细胞内ROS减少可降低氧化应激损伤,激活神经源性位点缺口同源蛋白(neurogenic loci gap homologous protein,Notch)信号通路,增加BMSCs成骨分化,促进骨形成[19].HIF-1α是调节线粒体代谢途径的关键转录因子,在TCA中下调HIF-1α会抑制骨细胞对葡萄糖的摄取和OC对骨吸收[20].因此,调节mtDNA数量和HIF-1α、PGC-1α基因表达,改变线粒体能量代谢途径,诱导BMSCs成骨分化可治疗OP. ...
1
... 细胞中产生ATP的方式主要有3种:一是糖酵解,二是TCA,三是线粒体OXPHOS.糖酵解和TCA产生还原型烟酰胺腺嘌呤二核苷酸、还原型黄素腺嘌呤二核苷酸和其他分子,而OXPHOS利用这些物质还原氧气并合成ATP[16].BMSCs增殖和自我更新时通过糖酵解获取能量,在分化时通过OXPHOS和TCA产生更多ATP,这可能是因为无氧代谢可避免ROS对mtDNA和细胞膜造成氧化损伤[17].在BMSCs成骨分化时线粒体膜蛋白和mtDNA数量显著增加,PGC-1α表达上调,促进OXPHOS产生ATP为其供能[18].抑制糖酵解时,线粒体呼吸减弱,ROS产生减少,而细胞内ROS减少可降低氧化应激损伤,激活神经源性位点缺口同源蛋白(neurogenic loci gap homologous protein,Notch)信号通路,增加BMSCs成骨分化,促进骨形成[19].HIF-1α是调节线粒体代谢途径的关键转录因子,在TCA中下调HIF-1α会抑制骨细胞对葡萄糖的摄取和OC对骨吸收[20].因此,调节mtDNA数量和HIF-1α、PGC-1α基因表达,改变线粒体能量代谢途径,诱导BMSCs成骨分化可治疗OP. ...
1
... 细胞中产生ATP的方式主要有3种:一是糖酵解,二是TCA,三是线粒体OXPHOS.糖酵解和TCA产生还原型烟酰胺腺嘌呤二核苷酸、还原型黄素腺嘌呤二核苷酸和其他分子,而OXPHOS利用这些物质还原氧气并合成ATP[16].BMSCs增殖和自我更新时通过糖酵解获取能量,在分化时通过OXPHOS和TCA产生更多ATP,这可能是因为无氧代谢可避免ROS对mtDNA和细胞膜造成氧化损伤[17].在BMSCs成骨分化时线粒体膜蛋白和mtDNA数量显著增加,PGC-1α表达上调,促进OXPHOS产生ATP为其供能[18].抑制糖酵解时,线粒体呼吸减弱,ROS产生减少,而细胞内ROS减少可降低氧化应激损伤,激活神经源性位点缺口同源蛋白(neurogenic loci gap homologous protein,Notch)信号通路,增加BMSCs成骨分化,促进骨形成[19].HIF-1α是调节线粒体代谢途径的关键转录因子,在TCA中下调HIF-1α会抑制骨细胞对葡萄糖的摄取和OC对骨吸收[20].因此,调节mtDNA数量和HIF-1α、PGC-1α基因表达,改变线粒体能量代谢途径,诱导BMSCs成骨分化可治疗OP. ...
1
... 细胞中产生ATP的方式主要有3种:一是糖酵解,二是TCA,三是线粒体OXPHOS.糖酵解和TCA产生还原型烟酰胺腺嘌呤二核苷酸、还原型黄素腺嘌呤二核苷酸和其他分子,而OXPHOS利用这些物质还原氧气并合成ATP[16].BMSCs增殖和自我更新时通过糖酵解获取能量,在分化时通过OXPHOS和TCA产生更多ATP,这可能是因为无氧代谢可避免ROS对mtDNA和细胞膜造成氧化损伤[17].在BMSCs成骨分化时线粒体膜蛋白和mtDNA数量显著增加,PGC-1α表达上调,促进OXPHOS产生ATP为其供能[18].抑制糖酵解时,线粒体呼吸减弱,ROS产生减少,而细胞内ROS减少可降低氧化应激损伤,激活神经源性位点缺口同源蛋白(neurogenic loci gap homologous protein,Notch)信号通路,增加BMSCs成骨分化,促进骨形成[19].HIF-1α是调节线粒体代谢途径的关键转录因子,在TCA中下调HIF-1α会抑制骨细胞对葡萄糖的摄取和OC对骨吸收[20].因此,调节mtDNA数量和HIF-1α、PGC-1α基因表达,改变线粒体能量代谢途径,诱导BMSCs成骨分化可治疗OP. ...



{kind=link}
{kind=link}