目的·探索弥漫性大B细胞淋巴瘤(diffuse large B cell lymphoma,DLBCL)中差异表达的性别决定区Y框转录因子9(SRY-box transcription factor 9,SOX9)基因所起到的作用,尤其是在生发中心B细胞(germinal center B-cell,GCB)来源亚型中对代谢重编程的调控作用。方法·选取NCICCR-DLBCL数据库中的481例DLBCL患者的临床信息和基因表达谱数据,使用R语言4.1.3版本进行数据分析与可视化,并基于RNA-seq测序表达量的细胞组织来源亚型(cell of origin subtype,COO)分类算法进行分类;使用ABC/GCB特征注释基因集,通过基因集富集分析(gene set enrichment analysis,GSEA)对分类进行验证。以SOX9的表达量高低将ABC和GCB亚组分别二分类。使用DEseq2包进行差异分析。使用KEGG(Kyoto Encyclopedia of Genes and Genomes)与Hallmark注释集分析SOX9与DLBCL的代谢的关系。采用Kaplan-Meier方法绘制生存曲线。采用GEPIA2进行泛癌分析。采用ESTIMATE包进行微环境评分分析。结果·481例DLBCL患者样本中,481例均有RNA-seq的表达量数据,421例有临床分期,335例有国际预后指数(international prognostic index,IPI)评分,234例有生存数据。分类得出ABC亚型232例(48.2%)、GCB亚型173例(36.0%)、未分类76例(15.8%),与数据库声明的比例相符,经富集分析验证符合ABC/GCB表达谱特征。SOX9低表达量组与SOX9高表达量组相比,总生存期更短,预后分数更差。泛癌分析示该现象亦可见于其他类型肿瘤。差异分析显示,在GCB亚型中,与SOX9高表达量组相比,SOX9低表达量组中有上调基因156个、下调基因1 826个。对于细胞代谢水平的变化,下调基因富集于糖酵解。结论·在ABC-DLBCL中,SOX9基因通过调控代谢重编程影响ABC-DLBCL的生物学特征。低表达SOX9的DLBCL,预示着肿瘤中糖酵解减少;其肿瘤基质细胞浸润程度更低,并且有着更差的预后。
关键词:性别决定区Y框转录因子9
;
弥漫性大B细胞淋巴瘤
;
代谢重编程
Abstract
Objective ·To explore the role played by the differentially expressed SRY-box transcription factor 9 (SOX9) gene in diffuse large B cell lymphoma (DLBCL), particularly in the regulation of metabolic reprogramming in the germinal center B-cell (GCB) like subtype. Methods ·The clinical information and gene expression profile data of 481 DLBCL patients retrieved from the NCICCR-DLBCL database were included. Data analysis and visualisation were performed by using R language version 4.1.3. The classification was performed by using a cell of origin subtype (COO) classification algorithm based on RNA-seq sequencing of expression. ABC/GCB features were used to annotate gene sets, and the classification was verified by gene set enrichment analysis. The ABC and GCB subgroup was dichotomised based on the mean expression of SOX9. Differential analysis was performed by using the DEseq2 package. The relationship between SOX9 and ABC-DLBCL metabolism was analysed by using KEGG (Kyoto Encyclopedia of Genes and Genomes) with the Hallmark annotation set. The survival curves were plotted by using the Kaplan-Meier method. The pan-cancer analysis was performed by using GEPIA2. The microenvironmental scoring analysis was performed by the ESTIMATE package. Results ·Of the 481 DLBCL patient samples, all the patients had RNA-seq expression data, 421 had clinical staging, 335 had international prognostic index (IPI) scores and 234 had survival data. The classification yielded 232 (48.2%) ABC subtypes, 173 (36.0%) GCB subtypes and 76 (15.8%) unclassified, consistent with the proportions declared in the database, and the enrichment analysis was verified to be consistent with the ABC/GCB expression profile. Compared to the high SOX9 expression group, the overall survival was shorter in the low SOX9 expression group and the prognostic score was worse. The pan-cancer analysis showed that this phenomenon was also seen in other tumor types. The differential analysis showed that there were 156 upregulated genes and 1 826 downregulated genes in the GCB subtype in the low SOX9 expression group, compared to the high SOX9 expression group. For metabolic processes, down-regulated genes were enriched in glycolysis. Conclusion ·In the ABC subtype of DLBCL, the SOX9 gene affects the biological features of ABC-DLBCL by regulating metabolic reprogramming, and low expression of SOX9 in DLBCL, possibly caused by high methylation, predicts decreased glycolysis in tumors. The proportion of tumor stromal cells decreases, showing a worse prognosis.
Keywords:SRY-box transcription factor 9 (SOX9)
;
diffuse large B cell lymphoma (DLBCL)
;
metabolic reprogramming
ZHANG Yirong, WEI Weiqing, MA Jiao, ZHANG Xue. Research on the role of SOX9 in regulating metabolic reprogramming in diffuse large B cell lymphoma. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(10): 1236-1244 doi:10.3969/j.issn.1674-8115.2023.10.003
弥漫性大B细胞淋巴瘤(diffuse large B cell lymphoma,DLBCL)是成人中最常见的淋巴瘤,具备很强的异质性,在形态学特点和生物学行为等方面均有多样化表现,对以蒽环类药物为基础的经典化学治疗(化疗)方案的预后差异明显[1]。故此前的研究基于基因表达特征定义了2种DLBCL亚型:生发中心B细胞(germinal center B-cell,GCB)亚型和活化B细胞(activated B-cell,ABC)亚型,GCB亚型的预后与对化疗的敏感度显著优于ABC亚型[1]。两者具备不同的遗传学基础、不同的分子生物学特征,故DLBCL的两种亚型也被视作不同的疾病,GCB淋巴瘤通常被认为起源于GCB,而ABC淋巴瘤起源于晚期GCB或浆母细胞的B细胞[2],但这些肿瘤的起源细胞在功能上尚未得到严格定义。有研究从表观遗传角度出发解释这两种亚型的起源,鉴定了一组基因[肿瘤坏死因子α(tumor necrosis factor-α,TNF-α)、性别决定区Y框转录因子9(SRY-box transcription factor 9,SOX9)等16个],它们在ABC和GCB的甲基化间存在显著差异并与其表达呈负相关,并且可以明确区分两种亚型[3]。因此了解上述基因在DLBCL发生发展中的功能与作用,将有助于理解DLBCL的发生发展过程,解释ABC与GCB亚型的起源与差异,对于明确此类患者发病机制及预后相关危险因素有着潜在临床意义,为靶向SOX9治疗DLBCL提供更多的可能性。
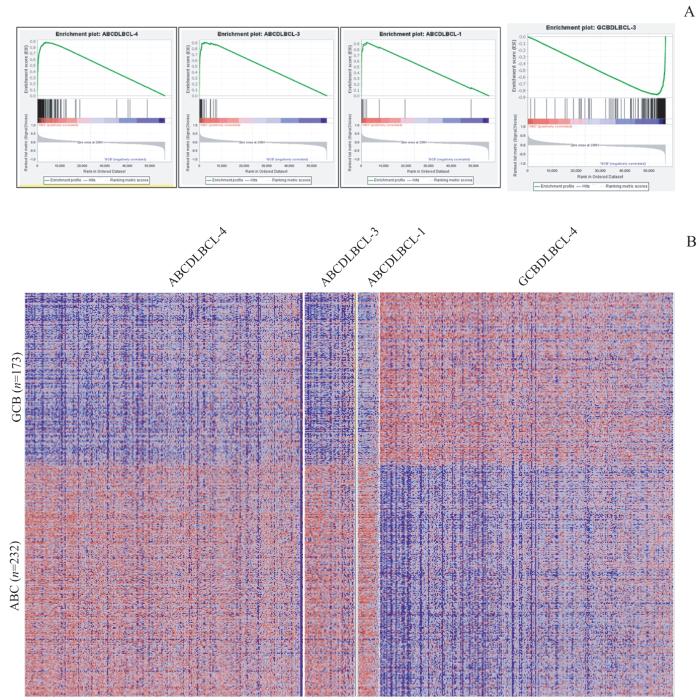
采用细胞组织来源亚型(cell of origin subtype,COO)分类算法进行ABC/GCB亚型分类。该算法由REDDY等[19]于2017年提出,基于RNA-seq所测得表达谱数据,以FPKM(fragments per kilobase of exon per million reads mapped)作为测量值,分位数归一化和log2转化的基因表达测量值在基因间进行z归一化。本研究通过分别取ABC、GCB亚型样本中基因表达z分数的平均值来计算每个样本的ABC分数和GCB分数,以ABC分数与GCB分数的差值作为综合亚型分数。如果综合亚型分数>0.25且其GCB分数<0.75,则样本被归类为ABC;如果综合亚型分数<-0.25且ABC分数<0.75,则将其归类为GCB。其余样本属于未分类组。
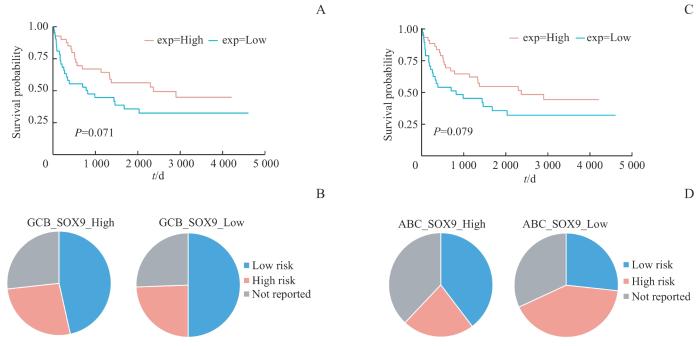
Note: A. Kaplan-Meier survival curve comparing the overall survival time for different SOX9 expression groups (upper 50% vs lower 50%) in GCB-DLBCL from NCICCR-DLBCL patient cohort. B. Distribution of IPI scores between different SOX9 groups (upper 50% vs lower 50%) in GCB-DLBCL from the NCICCR-DLBCL patient cohort. C. Kaplan-Meier survival curve comparing the overall survival time for different SOX9 expression groups (upper 50% vs lower 50%) in ABC-DLBCL from NCICCR-DLBCL patient cohort. D. Distribution of IPI scores between different SOX9 groups (upper 50% vs lower 50%) in ABC-DLBCL from the NCICCR-DLBCL patient cohort. "Low risk" contains "Low risk" and "Low-intermediate risk" in original cohort, likewise the "High". Statistical significance was determined by Fisher's exact test.
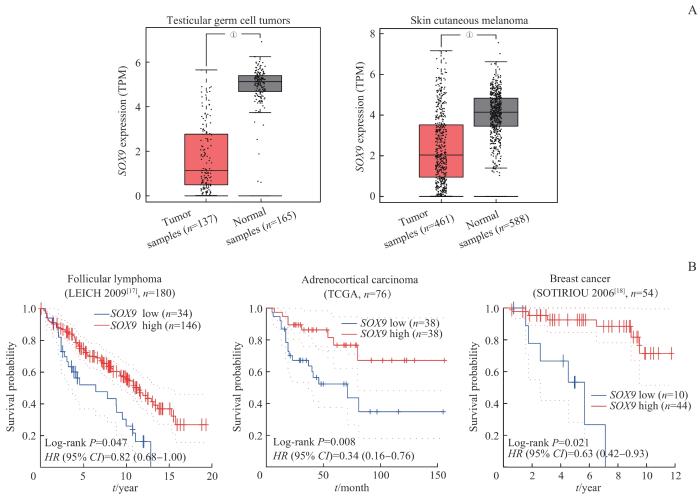
Note: A. Changes of SOX9 expression between tumor samples and normal samples from testicular germ cells tumors and skin cutaneous melanoma based on TCGA database. B. Kaplan-Meier survival curve comparing the overall survival time for different SOX9 expression groups in follicular lymphoma, adrenocortical carcinoma and breast cancer. Statistical significance was determined by the Log-rank test. ①P=0.000.
Fig 3
Expression and prognosis value of SOX9 in various cancer types
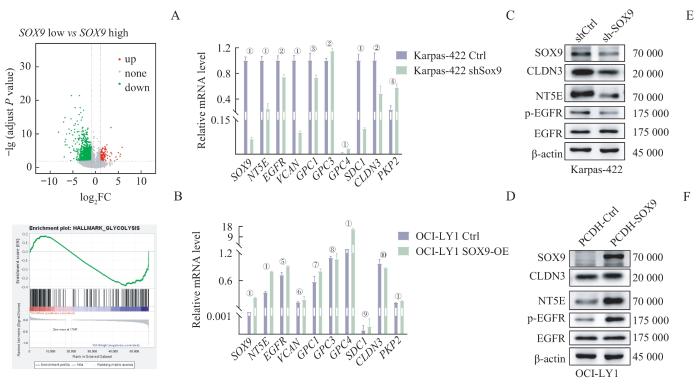
Note: A. Volcano plot of differential expression analysis. B. Representative GESA plots with hallmark and KEGG as reference gene sets. C/D. Change of glycolysis was confirmed by qPCR analysis in Karpas-422 (C) and OCI-LY1 (D) cell line. ①P=0.000, ②P=0.006, ③P=0.016, ④P=0.002, ⑤P=0.003, ⑥P=0.254, ⑦P=0.036, ⑧P=0.716, ⑨P=0.496, ⑩P=0.129. E/F. Change of glycolysis was confirmed by Western blotting analysis in Karpas-422 (E) and OCI-LY1 (F) cell line.
Fig 4
Regulation of metabolic pathways by SOX9 in GCB-DLBCL
ZHANG Yirong was responsible for conducting experiments, analyzing data and writing the manuscript. WEI Weiqing primarily handled data analysis. ZHANG Xue and MA Jiao contributed to the conception of the manuscript and provided guidance for its revision. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
All authors disclose no relevant conflict of interests.
ROSENWALD A, WRIGHT G, CHAN W C, et al. The use of molecular profiling to predict survival after chemotherapy for diffuse large-B-cell lymphoma[J]. N Engl J Med, 2002, 346(25): 1937-1947.
BEA S. Diffuse large B-cell lymphoma subgroups have distinct genetic profiles that influence tumor biology and improve gene-expression-based survival prediction[J]. Blood, 2005, 106(9): 3183-3190.
CONDELLI V, CRISPO F, PIETRAFESA M, et al. HSP90 molecular chaperones, metabolic rewiring, and epigenetics: impact on tumor progression and perspective for anticancer therapy[J]. Cells, 2019, 8(6): 532.
SUN N Y, YANG M H. Metabolic reprogramming and epithelial-mesenchymal plasticity: opportunities and challenges for cancer therapy[J]. Front Oncol, 2020, 10: 792.
YIN G, LIANG Y, WANG Y, et al. mTOR complex 1 signalling regulates the balance between lipid synthesis and oxidation in hypoxia lymphocytes[J]. Biosci Rep, 2017, 37(1): BSR20160479.
PANDA M, TRIPATHI S K, BISWAL B K. SOX9: an emerging driving factor from cancer progression to drug resistance[J]. Biochim Biophys Acta Rev Cancer, 2021, 1875(2): 188517.
ENJUANES A, FERNÀNDEZ V, HERNÁNDEZ L, et al. Identification of methylated genes associated with aggressive clinicopathological features in mantle cell lymphoma[J]. PLoS One, 2011, 6(5): e19736.
BENNETT L B, SCHNABEL J L, KELCHEN J M, et al. DNA hypermethylation accompanied by transcriptional repression in follicular lymphoma[J]. Genes Chromosom Cancer, 2009, 48(9): 828-841.
CHENG P F, SHAKHOVA O, WIDMER D S, et al. Methylation-dependent SOX9 expression mediates invasion in human melanoma cells and is a negative prognostic factor in advanced melanoma[J]. Genome Biol, 2015, 16(1): 42.
PASSERON T, VALENCIA J C, NAMIKI T, et al. Upregulation of SOX9 inhibits the growth of human and mouse melanomas and restores their sensitivity to retinoic acid[J]. J Clin Invest, 2009, 119(4): 954-963.
SLATTERY M L, HERRICK J S, MULLANY L E, et al. The co-regulatory networks of tumor suppressor genes, oncogenes, and miRNAs in colorectal cancer[J]. Genes Chromosom Cancer, 2017, 56(11): 769-787.
LEICH E, SALAVERRIA I, BEA S, et al. Follicular lymphomas with and without translocation t(14;18) differ in gene expression profiles and genetic alterations[J]. Blood, 2009, 114(4): 826-834.
SOTIRIOU C, WIRAPATI P, LOI S, et al. Gene expression profiling in breast cancer: understanding the molecular basis of histologic grade to improve prognosis[J]. J Natl Cancer Inst, 2006, 98(4): 262-272.
SHAFFER A L, WRIGHT G, YANG L M, et al. A library of gene expression signatures to illuminate normal and pathological lymphoid biology[J]. Immunol Rev, 2006, 210: 67-85.
SUBRAMANIAN A, TAMAYO P, MOOTHA V K, et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles[J]. Proc Natl Acad Sci USA, 2005, 102(43): 15545-15550.
YOSHIHARA K, SHAHMORADGOLI M, MARTÍNEZ E, et al. Inferring tumour purity and stromal and immune cell admixture from expression data[J]. Nat Commun, 2013, 4: 2612.
JIANG Y W, HATZI K, ELEMENTO O, et al. Enhancer profiling reveals SOX9 as a novel transcription regulator of B cell activation and DLBCL transformation[J]. Blood, 2012, 120(21): 527.
SHEN Y J, ZHOU J Q, NIE K, et al. Oncogenic role of the SOX9-DHCR24-cholesterol biosynthesis axis in IGH-BCL2+ diffuse large B-cell lymphomas[J]. Blood, 2022, 139(1): 73-86.
GOOPTU M, WHITAKER-MENEZES D, SPRANDIO J, et al. Mitochondrial and glycolytic metabolic compartmentalization in diffuse large B-cell lymphoma[J]. Semin Oncol, 2017, 44(3): 204-217.
LAM L T, WRIGHT G, DAVIS R E, et al. Cooperative signaling through the signal transducer and activator of transcription 3 and nuclear factor-κB pathways in subtypes of diffuse large B-cell lymphoma[J]. Blood, 2008, 111(7): 3701-3713.
... 弥漫性大B细胞淋巴瘤(diffuse large B cell lymphoma,DLBCL)是成人中最常见的淋巴瘤,具备很强的异质性,在形态学特点和生物学行为等方面均有多样化表现,对以蒽环类药物为基础的经典化学治疗(化疗)方案的预后差异明显[1].故此前的研究基于基因表达特征定义了2种DLBCL亚型:生发中心B细胞(germinal center B-cell,GCB)亚型和活化B细胞(activated B-cell,ABC)亚型,GCB亚型的预后与对化疗的敏感度显著优于ABC亚型[1].两者具备不同的遗传学基础、不同的分子生物学特征,故DLBCL的两种亚型也被视作不同的疾病,GCB淋巴瘤通常被认为起源于GCB,而ABC淋巴瘤起源于晚期GCB或浆母细胞的B细胞[2],但这些肿瘤的起源细胞在功能上尚未得到严格定义.有研究从表观遗传角度出发解释这两种亚型的起源,鉴定了一组基因[肿瘤坏死因子α(tumor necrosis factor-α,TNF-α)、性别决定区Y框转录因子9(SRY-box transcription factor 9,SOX9)等16个],它们在ABC和GCB的甲基化间存在显著差异并与其表达呈负相关,并且可以明确区分两种亚型[3].因此了解上述基因在DLBCL发生发展中的功能与作用,将有助于理解DLBCL的发生发展过程,解释ABC与GCB亚型的起源与差异,对于明确此类患者发病机制及预后相关危险因素有着潜在临床意义,为靶向SOX9治疗DLBCL提供更多的可能性. ...
... 弥漫性大B细胞淋巴瘤(diffuse large B cell lymphoma,DLBCL)是成人中最常见的淋巴瘤,具备很强的异质性,在形态学特点和生物学行为等方面均有多样化表现,对以蒽环类药物为基础的经典化学治疗(化疗)方案的预后差异明显[1].故此前的研究基于基因表达特征定义了2种DLBCL亚型:生发中心B细胞(germinal center B-cell,GCB)亚型和活化B细胞(activated B-cell,ABC)亚型,GCB亚型的预后与对化疗的敏感度显著优于ABC亚型[1].两者具备不同的遗传学基础、不同的分子生物学特征,故DLBCL的两种亚型也被视作不同的疾病,GCB淋巴瘤通常被认为起源于GCB,而ABC淋巴瘤起源于晚期GCB或浆母细胞的B细胞[2],但这些肿瘤的起源细胞在功能上尚未得到严格定义.有研究从表观遗传角度出发解释这两种亚型的起源,鉴定了一组基因[肿瘤坏死因子α(tumor necrosis factor-α,TNF-α)、性别决定区Y框转录因子9(SRY-box transcription factor 9,SOX9)等16个],它们在ABC和GCB的甲基化间存在显著差异并与其表达呈负相关,并且可以明确区分两种亚型[3].因此了解上述基因在DLBCL发生发展中的功能与作用,将有助于理解DLBCL的发生发展过程,解释ABC与GCB亚型的起源与差异,对于明确此类患者发病机制及预后相关危险因素有着潜在临床意义,为靶向SOX9治疗DLBCL提供更多的可能性. ...
3
... 弥漫性大B细胞淋巴瘤(diffuse large B cell lymphoma,DLBCL)是成人中最常见的淋巴瘤,具备很强的异质性,在形态学特点和生物学行为等方面均有多样化表现,对以蒽环类药物为基础的经典化学治疗(化疗)方案的预后差异明显[1].故此前的研究基于基因表达特征定义了2种DLBCL亚型:生发中心B细胞(germinal center B-cell,GCB)亚型和活化B细胞(activated B-cell,ABC)亚型,GCB亚型的预后与对化疗的敏感度显著优于ABC亚型[1].两者具备不同的遗传学基础、不同的分子生物学特征,故DLBCL的两种亚型也被视作不同的疾病,GCB淋巴瘤通常被认为起源于GCB,而ABC淋巴瘤起源于晚期GCB或浆母细胞的B细胞[2],但这些肿瘤的起源细胞在功能上尚未得到严格定义.有研究从表观遗传角度出发解释这两种亚型的起源,鉴定了一组基因[肿瘤坏死因子α(tumor necrosis factor-α,TNF-α)、性别决定区Y框转录因子9(SRY-box transcription factor 9,SOX9)等16个],它们在ABC和GCB的甲基化间存在显著差异并与其表达呈负相关,并且可以明确区分两种亚型[3].因此了解上述基因在DLBCL发生发展中的功能与作用,将有助于理解DLBCL的发生发展过程,解释ABC与GCB亚型的起源与差异,对于明确此类患者发病机制及预后相关危险因素有着潜在临床意义,为靶向SOX9治疗DLBCL提供更多的可能性. ...
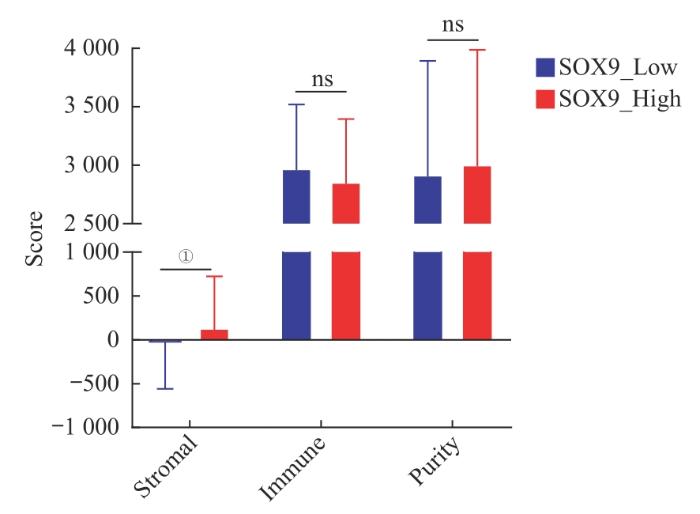
... 基于HPA数据库中的IHC研究,绝大多数原发性非霍奇金性淋巴瘤组织中检测不到SOX9的表达.在癌症细胞系百科全书(Cancer Cell Line Encyclopedia,CCLE)数据库中对各细胞系表达量的测定也显示,绝大多数DLBCL细胞株的SOX9表达量极低.此前的研究鉴定了低表达SOX9是ABC亚型的特征[3],而ABC亚型往往意味着更差的预后.此外有研究[23]发现,在DLBCL细胞中敲减SOX9将提高肿瘤组织的集落形成能力.这些观察提示SOX9的表达缺失可能促进DLBCL的发生发展,不利于患者的预后.尽管我们此前的研究表明了SOX9高表达在IGH-BCL2易位突变的DLBCL亚型中通过调控24-脱氢胆固醇还原酶(dehydrocholesterol reductase 24,DHCR24)的表达,促进GCB亚型DLBCL细胞的胆固醇代谢,从而诱导DLBCL的发生[24],但我们的结果显示在DLBCL中,SOX9高表达的现象仅出现在GCB亚型的两株DLBCL细胞系中(Karpas-422和DB细胞).在这类特定亚型中的SOX9可能通过特有机制发挥致癌效应.总之,在绝大多数的DLBCL中SOX9表达缺失的生物学效应仍有待阐述,我们的数据揭示了以前未描述的SOX9表达缺失的致癌和预后价值. ...
... 采用细胞组织来源亚型(cell of origin subtype,COO)分类算法进行ABC/GCB亚型分类.该算法由REDDY等[19]于2017年提出,基于RNA-seq所测得表达谱数据,以FPKM(fragments per kilobase of exon per million reads mapped)作为测量值,分位数归一化和log2转化的基因表达测量值在基因间进行z归一化.本研究通过分别取ABC、GCB亚型样本中基因表达z分数的平均值来计算每个样本的ABC分数和GCB分数,以ABC分数与GCB分数的差值作为综合亚型分数.如果综合亚型分数>0.25且其GCB分数<0.75,则样本被归类为ABC;如果综合亚型分数<-0.25且ABC分数<0.75,则将其归类为GCB.其余样本属于未分类组. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}