目的·研究gasdermin D(GSDMD)抑制剂necrosulfonamide(NSA)通过抑制肺上皮细胞焦亡对脂多糖(lipopolysaccharide,LPS)诱导的新生大鼠支气管肺发育不良(bronchopulmonary dysplasia,BPD)肺泡化阻滞的影响。方法·将孕SD大鼠随机分为对照组、BPD组、BPD+NSA组和NSA组,羊膜腔注射LPS建立新生鼠BPD模型。取各组出生后第1、3、7日新生鼠肺组织,通过苏木精-伊红(H-E)染色观察肺泡化情况;利用免疫荧光法检测各组新生鼠肺部GSDMD-N端蛋白表达情况;荧光定量PCR法检测新生鼠肺组织中炎症因子白介素-1β(interleukin-1β,IL-1β)mRNA水平。体外培养小鼠肺泡上皮细胞系MLE-12,给予LPS以及腺苷三磷酸(adenosine triphosphate,ATP)刺激和NSA干预,CCK-8法检测MLE-12细胞活力,Hoechst 33342和碘化丙啶(propidium iodide,PI)染色法检测细胞焦亡水平,免疫荧光法检测MLE-12细胞表面活性剂蛋白C(surfactant protein C,SFTPC)和GSDMD-N端蛋白的表达情况。结果·体内实验结果显示:羊膜腔内注射LPS可导致肺发育受阻,模拟BPD的病理改变;羊膜腔内注射LPS建立的BPD模型中肺泡上皮细胞GSDMD-N端表达升高;NSA干预明显改善了BPD新生鼠的肺发育阻滞并抑制了IL-1β的mRNA表达(均P<0.05)。体外实验结果显示:LPS/ATP刺激下肺泡上皮细胞MLE-12活力下降,发生焦亡;NSA干预提高了肺泡上皮细胞活力并且抑制了焦亡(均P<0.05);NSA上调了LPS/ATP刺激下肺泡上皮细胞SFTPC的表达,抑制了LPS/ATP刺激下肺泡上皮细胞GSDMD-N端表达的上调(均P<0.05)。结论·抑制肺泡上皮细胞焦亡可改善LPS诱导的BPD新生鼠肺组织病理学改变。
关键词:支气管肺发育不良
;
细胞焦亡
;
gasdermin D
;
肺泡上皮细胞
Abstract
Objective ·To study the effect of gasdermin D (GSDMD) inhibitor necrosulfonamide (NSA) on alveolarization arrest in lipopolysaccharide (LPS)-induced bronchopulmonary dysplasia (BPD) newborn rats via inhibiting alveolar epithelial cell pyroptosis. Methods ·Pregnant SD rats were randomly assigned to four groups as follows: control, BPD, BPD with NSA and NSA group, and then were prepared to receive intra-amniotic injection of LPS. Lung tissues of newborn rats on the first, third and seventh day after birth were stained by hematoxylin-eosin (H-E) to observe lung development. The expressions of GSDMD-N-terminal in lungs of newborn rats in each group were detected by immunofluorescence. The mRNA levels of interleukin-1β (IL-1β) of newborn rats' lungs was detected by real-time PCR. In vitro, the mouse alveolar epithelial cell line MLE-12 was cultured and treated with LPS/adenosine triphosphate (ATP) and NSA. The cell viability of MLE-12 cells was detected by CCK-8 method, the pyroptosis was detected by Hoechst 33342 and propidium iodide (PI) staining, and the expressions of surfactant protein C (SFTPC) and GSDMD-N protein in MLE-12 cells were detected by immunofluorescence. Results ·In vivo, intra-amniotic injection of LPS hindered lung development, resulting in the pathological hallmarks of BPD. The GSDMD-N expression of alveolar epithelial cells increased in the BPD rat model established by intra-amniotic injection of LPS, while NSA treatment significantly improved the lung development of BPD rats and inhibited the IL-1β mRNA expression (both P<0.05). In vitro, the study confirmed that LPS/ATP treatment decreased the viability of alveolar epithelial cells MLE-12 and induced pyroptosis, while NSA treatment increased alveolar epithelial cell viability and inhibited pyroptosis (both P<0.05). In addition, NSA treatment upregulated the SFTPC expression and inhibited the GSDMD-N expression in LPS/ATP-stimulated alveolar epithelial cells (both P<0.05). Conclusion ·Inhibiting the alveolar epithelial cell pyroptosis can improve the alveolar development in BPD newborn rats.
ZHENG Xiaoyan, WANG Xingyun, ZHANG Yongjun. Improvement of alveolarization arrest in newborn rats with bronchopulmonary dysplasia via inhibiting alveolar epithelial cell pyroptosis. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(2): 171-179 doi:10.3969/j.issn.1674-8115.2023.02.005
细胞焦亡是一种炎性含半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,caspase)依赖性的细胞死亡,其特征是细胞肿胀、溶解和多种炎症因子的释放。多种炎症小体的激活可活化caspase。活性caspase一方面切割焦亡效应蛋白gasdermin D(GSDMD)形成GSDMD-N端,而GSDMD-N端在细胞膜上寡聚形成微孔,导致细胞渗透压的变化,进而发生肿胀直至细胞膜破裂;另一方面裂解白介素-1β(interleukin-1β,IL-1β)前体使其成为成熟的IL-1β,并通过GSDMD小孔释放从而引发过度炎症反应,造成组织损伤[6-7]。细胞焦亡在BPD的发生和发展中起到的重要作用已经被许多研究证实。NOD样受体热蛋白结构域相关蛋白3(NOD-like receptor thermal protein domain associated protein 3,NLRP3)炎症小体及NLRP1炎症小体被认为在BPD的发生和发展中发挥着重要作用[8-9]。此外,抑制caspase-1活性或IL-1β分泌均可改善BPD新生鼠的肺损伤[9-10]。而作为肺发育主体的肺泡上皮细胞,其焦亡引发的呼吸系统疾病也被广泛报道[11-12]。Necrosulfonamide(NSA)是一种GSDMD特异性抑制剂,不仅能抑制GSDMD蛋白的切割,还能抑制GSDMD-N端寡聚化和膜穿孔的形成[13-14]。但抑制焦亡效应蛋白GSDMD是否可以抑制肺泡上皮细胞的焦亡从而改善BPD,目前尚不清楚。因此,本研究拟探究GSDMD抑制剂NSA对羊膜腔内注射脂多糖(lipopolysaccharide,LPS)诱导的BPD新生大鼠肺发育的影响以及对其肺泡上皮细胞焦亡的抑制作用。
1 材料与方法
1.1 材料
1.1.1 实验动物与细胞来源
孕16 d SPF级Sprague Dawley(SD)大鼠购自上海吉辉实验动物有限责任公司,动物生产许可证号SCXK(沪)2017-0012。实验动物饲养及实验操作于上海交通大学医学院附属新华医院实验动物中心进行,动物使用许可证号SYXK9(沪)2013-0106。小鼠肺泡上皮细胞(MLE-12细胞)购自中国科学院上海生命科学研究院细胞资源中心。
1.1.2 主要试剂
LPS、腺苷三磷酸(adenosine triphosphate,ATP)和NSA均购于美国MedChemExpress公司;DMEM/F12培养基、胎牛血清(fetal bovine serum,FBS)、青链霉素双抗溶液(penicillin-streptomycin,PS)均购自美国Gibco公司;兔多克隆GSDMD-N端抗体购自江苏亲科生物研究中心有限公司;抗表面活性剂蛋白C(surfactant protein C,SFTPC)抗体购自美国Proteintech公司;CCK-8试剂盒、Hoechst 33342荧光染料/碘化丙啶(propidium iodide,PI)双染试剂盒和快速封闭液均购自上海碧云天生物技术有限公司。
待孕鼠分娩后,记当日为新生鼠出生后第0日(postnatal day 0,P0),待出生后1、3、7 d采集新生鼠肺组织样本。腹腔麻醉后,打开新生鼠胸腔暴露肺部,用冰PBS灌洗肺血管,用缝合线结扎右肺根部和气管,用4%多聚甲醛灌注左侧肺叶,直至肺尖边缘轻微隆起。摘除左肺上叶将其浸泡在4%多聚甲醛中进行固定,右肺放入EP管中迅速置于液氮中冻存,-80 ℃保存。
1.2.3 苏木精-伊红染色观察肺组织病理改变
左肺固定24 h后,经过脱水、浸蜡、包埋、切片、苏木精-伊红(H-E)染色、封片后,置于显微镜下观察。各组随机选取5个样本,每个样本取最大肺部横截面的3个不同层面的切片,每张切片随机选取不重叠的3个视野拍照保存。采用图像处理软件Image J进行图像处理和肺组织形态学分析的计数,统计指标包括平均肺泡数、平均次级间隔数、平均肺泡内衬间隔(mean linear intercept,MLI)。MLI的计算方法如下:经切片视野中心点画十字交叉线,测出总长度(L),计数线上的所有肺泡数(N),MLI = L/N(单位:µm)。
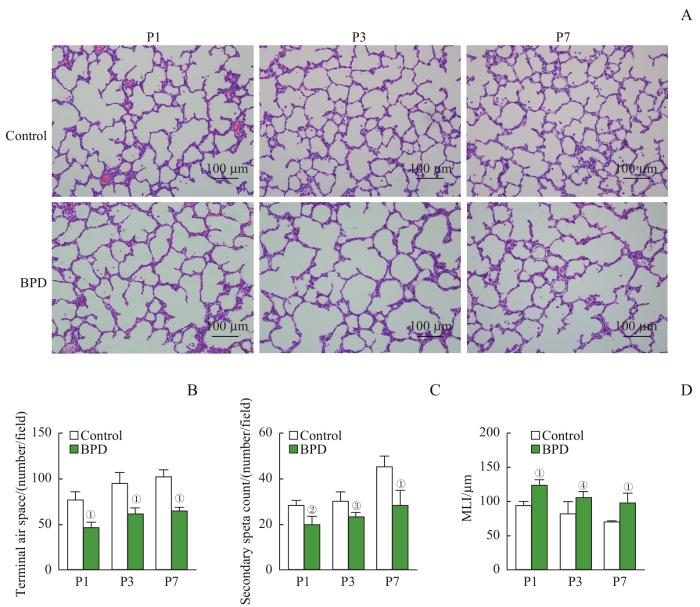
Note: A. The representative lung sections stained with H-E staining. B. The quantification of the terminal air spaces of lung tissues from newborn rats. C. The quantification of the secondary septa of lung tissues from newborn rats. D. The quantification of the MLI of lung tissues from newborn rats. ①P=0.000, ②P=0.010, ③P=0.038, ④P=0.004, compared with the control group.
Fig 1
Alveolar developmental arrest caused by intra-amniotic LPS injection
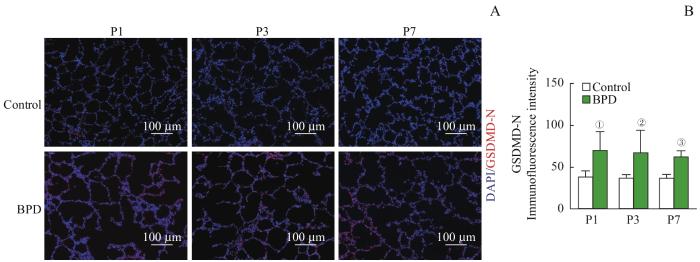
Note:A. Representative immunofluorescence images of GSDMD-N terminal (red) in the lungs of newborn rats. B. Quantitation of GSDMD-N terminal fluorescence intensity. ①P=0.007, ②P=0.015, ③P=0.033, compared with the control group.
Fig 2
Alveolar epithelial cell pyroptosis in BPD newborn rats
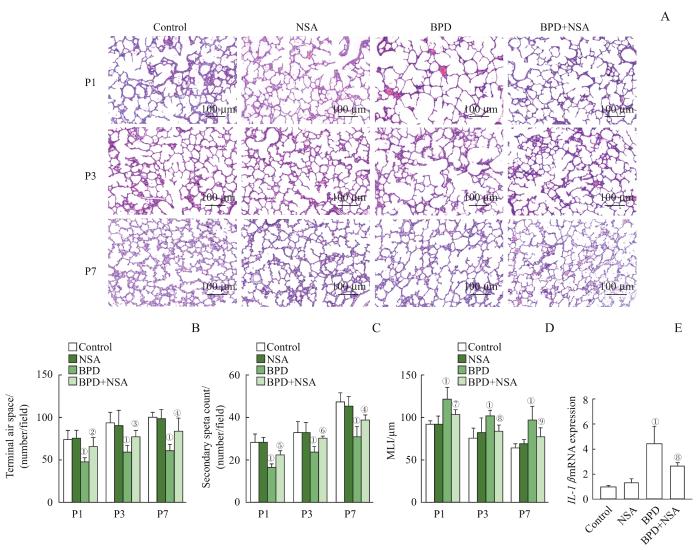
Note:A. The representative lung sections stained with H-E staining. B. The quantification of the terminal air spaces of lung tissues from newborn rats. C. The quantification of the secondary septa of lung tissues from newborn rats. D. The quantification of the MLI of lung tissues from newborn rats. E. Comparison of IL-1β mRNA among the groups. ①P=0.000, compared with the control group; ②P=0.031, ③P=0.049, ④P=0.002, ⑤P=0.040, ⑥P=0.020, ⑦P=0.024, ⑧P =0.026, ⑨P =0.014, compared with the BPD group.
Fig 3
Effects of NSA on lung pathology and expression of IL-1β mRNA in newborn rats
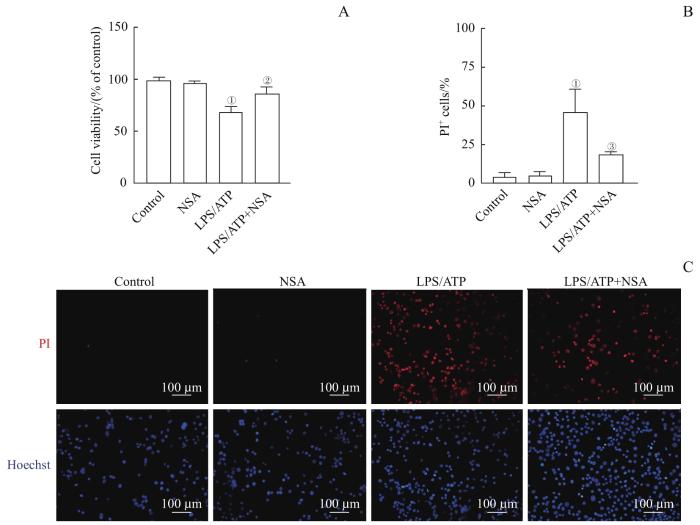
Note:A. Cell viability of MLE-12 detected by CCK-8 assay. B. Percentages of PI+ cells. C. Pyroptotic cells detected by Hoechst 33342 and PI staining. ①P=0.000, compared with the control group; ②P=0.003, ③P=0.008, compared with the LPS/ATP group.
Fig 4
Inhibition effect of NSA on LPS/ATP-induced pyroptosis in MLE-12 cells
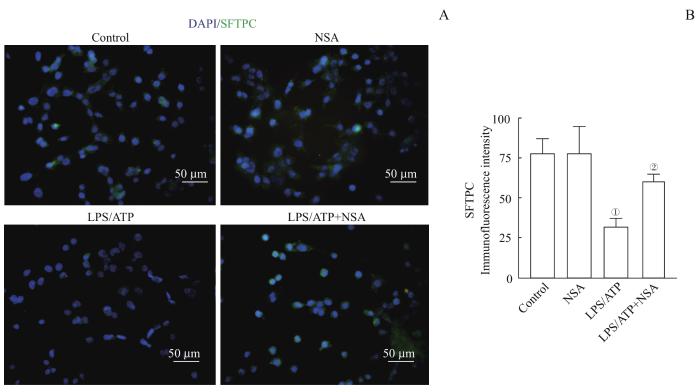
Note:A. Representative immunofluorescence images of SFTPC (green) in MLE-12. B. Quantitation of SFTPC fluorescence intensity. ①P=0.002, compared with the control group; ②P=0.035, compared with the LPS/ATP group.
Fig 5
Effect of NSA treatment on SFTPC expression in MLE-12 cells
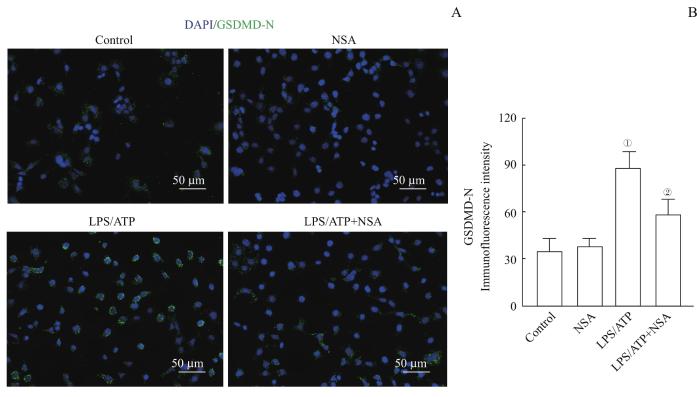
Note:A. Representative immunofluorescence images of GSDMD-N terminal (green) in MLE-12. B. Quantitation of GSDMD-N terminal fluorescence intensity. ①P=0.000, compared with the control group; ②P=0.010, compared with the LPS/ATP group.
Fig 6
Effect of NSA treatment on GSDMD-N expression in MLE-12 cells
The study was designed by ZHENG Xiaoyan, WANG Xingyun and ZHANG Yongjun. The research implementation and data analysis were conducted by ZHENG Xiaoyan. The manuscript was drafted and revised by ZHENG Xiaoyan, WANG Xingyun and ZHANG Yongjun. All authors have read the last version and revised and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
STOLL B J, HANSEN N I, BELL E F, et al. Trends in care practices, morbidity, and mortality of extremely preterm neonates, 1993-2012[J]. JAMA, 2015, 314(10): 1039-1051.
BELL E F, HINTZ S R, HANSEN N I, et al. Mortality, in-hospital morbidity, care practices, and 2-year outcomes for extremely preterm infants in the US, 2013-2018[J]. JAMA, 2022, 327(3): 248-263.
ISLAM J Y, KELLER R L, ASCHNER J L, et al. Understanding the short- and long-term respiratory outcomes of prematurity and bronchopulmonary dysplasia[J]. Am J Respir Crit Care Med, 2015, 192(2): 134-156.
LIAO J, KAPADIA V S, BROWN L S, et al. The NLRP3 inflammasome is critically involved in the development of bronchopulmonary dysplasia[J]. Nat Commun, 2015, 6: 8977.
STOUCH A N, MCCOY A M, GREER R M, et al. IL-1β and inflammasome activity link inflammation to abnormal fetal airway development[J]. J Immunol, 2016, 196(8): 3411-3420.
KONG X, GAO M, LIU Y, et al. GSDMD-miR-223-NLRP3 axis involved in B(a)P-induced inflammatory injury of alveolar epithelial cells[J]. Ecotoxicol Environ Saf, 2022, 232: 113286.
WAN X P, LI J Q, WANG Y P, et al. H7N9 virus infection triggers lethal cytokine storm by activating gasdermin E-mediated pyroptosis of lung alveolar epithelial cells[J]. Natl Sci Rev, 2022, 9(1): nwab137.
RATHKEY J K, ZHAO J, LIU Z, et al. Chemical disruption of the pyroptotic pore-forming protein gasdermin D inhibits inflammatory cell death and sepsis[J]. Sci Immunol, 2018, 3(26): eaat2738.
KALIKKOT THEKKEVEEDU R, GUAMAN M C, SHIVANNA B. Bronchopulmonary dysplasia: a review of pathogenesis and pathophysiology[J]. Respir Med, 2017, 132: 170-177.
ALI A, ZAMBRANO R, DUNCAN M R, et al. Hyperoxia-activated circulating extracellular vesicles induce lung and brain injury in neonatal rats[J]. Sci Rep, 2021, 11(1): 8791.
CAYABYAB R G, JONES C A, KWONG K Y, et al. Interleukin-1β in the bronchoalveolar lavage fluid of premature neonates: a marker for maternal chorioamnionitis and predictor of adverse neonatal outcome[J]. J Matern Fetal Neonatal Med, 2003, 14(3): 205-211.
ZHANG Q, RAN X, HE Y, et al. Acetate downregulates the activation of NLRP3 inflammasomes and attenuates lung injury in neonatal mice with bronchopulmonary dysplasia[J]. Front Pediatr, 2021, 8: 595157.
LU F, LAN Z, XIN Z, et al. Emerging insights into molecular mechanisms underlying pyroptosis and functions of inflammasomes in diseases[J]. J Cell Physiol, 2020, 235(4): 3207-3221.
DHANI S, ZHAO Y, ZHIVOTOVSKY B. A long way to go: caspase inhibitors in clinical use[J]. Cell Death Dis, 2021, 12(10): 949.
BIALER M, JOHANNESSEN S I, KUPFERBERG H J, et al. Progress report on new antiepileptic drugs: a summary of the Eleventh Eilat Conference (EILAT Ⅺ)[J]. Epilepsy Res, 2013, 103(1): 2-30.
FLORES J, NOËL A, FOVEAU B, et al. Pre-symptomatic caspase-1 inhibitor delays cognitive decline in a mouse model of Alzheimer disease and aging[J]. Nat Commun, 2020, 11(1): 4571.
HIRANI D, ALVIRA C M, DANOPOULOS S, et al. Macrophage-derived IL-6 trans-signalling as a novel target in the pathogenesis of bronchopulmonary dysplasia[J]. Eur Respir J, 2022, 59(2): 2002248.
HOU A, FU J, YANG H, et al. Hyperoxia stimulates the transdifferentiation of type Ⅱ alveolar epithelial cells in newborn rats[J]. Am J Physiol Lung Cell Mol Physiol, 2015, 308(9): L861-L872.
... 细胞焦亡是一种炎性含半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,caspase)依赖性的细胞死亡,其特征是细胞肿胀、溶解和多种炎症因子的释放.多种炎症小体的激活可活化caspase.活性caspase一方面切割焦亡效应蛋白gasdermin D(GSDMD)形成GSDMD-N端,而GSDMD-N端在细胞膜上寡聚形成微孔,导致细胞渗透压的变化,进而发生肿胀直至细胞膜破裂;另一方面裂解白介素-1β(interleukin-1β,IL-1β)前体使其成为成熟的IL-1β,并通过GSDMD小孔释放从而引发过度炎症反应,造成组织损伤[6-7].细胞焦亡在BPD的发生和发展中起到的重要作用已经被许多研究证实.NOD样受体热蛋白结构域相关蛋白3(NOD-like receptor thermal protein domain associated protein 3,NLRP3)炎症小体及NLRP1炎症小体被认为在BPD的发生和发展中发挥着重要作用[8-9].此外,抑制caspase-1活性或IL-1β分泌均可改善BPD新生鼠的肺损伤[9-10].而作为肺发育主体的肺泡上皮细胞,其焦亡引发的呼吸系统疾病也被广泛报道[11-12].Necrosulfonamide(NSA)是一种GSDMD特异性抑制剂,不仅能抑制GSDMD蛋白的切割,还能抑制GSDMD-N端寡聚化和膜穿孔的形成[13-14].但抑制焦亡效应蛋白GSDMD是否可以抑制肺泡上皮细胞的焦亡从而改善BPD,目前尚不清楚.因此,本研究拟探究GSDMD抑制剂NSA对羊膜腔内注射脂多糖(lipopolysaccharide,LPS)诱导的BPD新生大鼠肺发育的影响以及对其肺泡上皮细胞焦亡的抑制作用. ...
2
... 细胞焦亡是一种炎性含半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,caspase)依赖性的细胞死亡,其特征是细胞肿胀、溶解和多种炎症因子的释放.多种炎症小体的激活可活化caspase.活性caspase一方面切割焦亡效应蛋白gasdermin D(GSDMD)形成GSDMD-N端,而GSDMD-N端在细胞膜上寡聚形成微孔,导致细胞渗透压的变化,进而发生肿胀直至细胞膜破裂;另一方面裂解白介素-1β(interleukin-1β,IL-1β)前体使其成为成熟的IL-1β,并通过GSDMD小孔释放从而引发过度炎症反应,造成组织损伤[6-7].细胞焦亡在BPD的发生和发展中起到的重要作用已经被许多研究证实.NOD样受体热蛋白结构域相关蛋白3(NOD-like receptor thermal protein domain associated protein 3,NLRP3)炎症小体及NLRP1炎症小体被认为在BPD的发生和发展中发挥着重要作用[8-9].此外,抑制caspase-1活性或IL-1β分泌均可改善BPD新生鼠的肺损伤[9-10].而作为肺发育主体的肺泡上皮细胞,其焦亡引发的呼吸系统疾病也被广泛报道[11-12].Necrosulfonamide(NSA)是一种GSDMD特异性抑制剂,不仅能抑制GSDMD蛋白的切割,还能抑制GSDMD-N端寡聚化和膜穿孔的形成[13-14].但抑制焦亡效应蛋白GSDMD是否可以抑制肺泡上皮细胞的焦亡从而改善BPD,目前尚不清楚.因此,本研究拟探究GSDMD抑制剂NSA对羊膜腔内注射脂多糖(lipopolysaccharide,LPS)诱导的BPD新生大鼠肺发育的影响以及对其肺泡上皮细胞焦亡的抑制作用. ...
... 细胞焦亡是一种炎性含半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,caspase)依赖性的细胞死亡,其特征是细胞肿胀、溶解和多种炎症因子的释放.多种炎症小体的激活可活化caspase.活性caspase一方面切割焦亡效应蛋白gasdermin D(GSDMD)形成GSDMD-N端,而GSDMD-N端在细胞膜上寡聚形成微孔,导致细胞渗透压的变化,进而发生肿胀直至细胞膜破裂;另一方面裂解白介素-1β(interleukin-1β,IL-1β)前体使其成为成熟的IL-1β,并通过GSDMD小孔释放从而引发过度炎症反应,造成组织损伤[6-7].细胞焦亡在BPD的发生和发展中起到的重要作用已经被许多研究证实.NOD样受体热蛋白结构域相关蛋白3(NOD-like receptor thermal protein domain associated protein 3,NLRP3)炎症小体及NLRP1炎症小体被认为在BPD的发生和发展中发挥着重要作用[8-9].此外,抑制caspase-1活性或IL-1β分泌均可改善BPD新生鼠的肺损伤[9-10].而作为肺发育主体的肺泡上皮细胞,其焦亡引发的呼吸系统疾病也被广泛报道[11-12].Necrosulfonamide(NSA)是一种GSDMD特异性抑制剂,不仅能抑制GSDMD蛋白的切割,还能抑制GSDMD-N端寡聚化和膜穿孔的形成[13-14].但抑制焦亡效应蛋白GSDMD是否可以抑制肺泡上皮细胞的焦亡从而改善BPD,目前尚不清楚.因此,本研究拟探究GSDMD抑制剂NSA对羊膜腔内注射脂多糖(lipopolysaccharide,LPS)诱导的BPD新生大鼠肺发育的影响以及对其肺泡上皮细胞焦亡的抑制作用. ...
... 细胞焦亡是一种炎性含半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,caspase)依赖性的细胞死亡,其特征是细胞肿胀、溶解和多种炎症因子的释放.多种炎症小体的激活可活化caspase.活性caspase一方面切割焦亡效应蛋白gasdermin D(GSDMD)形成GSDMD-N端,而GSDMD-N端在细胞膜上寡聚形成微孔,导致细胞渗透压的变化,进而发生肿胀直至细胞膜破裂;另一方面裂解白介素-1β(interleukin-1β,IL-1β)前体使其成为成熟的IL-1β,并通过GSDMD小孔释放从而引发过度炎症反应,造成组织损伤[6-7].细胞焦亡在BPD的发生和发展中起到的重要作用已经被许多研究证实.NOD样受体热蛋白结构域相关蛋白3(NOD-like receptor thermal protein domain associated protein 3,NLRP3)炎症小体及NLRP1炎症小体被认为在BPD的发生和发展中发挥着重要作用[8-9].此外,抑制caspase-1活性或IL-1β分泌均可改善BPD新生鼠的肺损伤[9-10].而作为肺发育主体的肺泡上皮细胞,其焦亡引发的呼吸系统疾病也被广泛报道[11-12].Necrosulfonamide(NSA)是一种GSDMD特异性抑制剂,不仅能抑制GSDMD蛋白的切割,还能抑制GSDMD-N端寡聚化和膜穿孔的形成[13-14].但抑制焦亡效应蛋白GSDMD是否可以抑制肺泡上皮细胞的焦亡从而改善BPD,目前尚不清楚.因此,本研究拟探究GSDMD抑制剂NSA对羊膜腔内注射脂多糖(lipopolysaccharide,LPS)诱导的BPD新生大鼠肺发育的影响以及对其肺泡上皮细胞焦亡的抑制作用. ...
... 细胞焦亡是一种炎性含半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,caspase)依赖性的细胞死亡,其特征是细胞肿胀、溶解和多种炎症因子的释放.多种炎症小体的激活可活化caspase.活性caspase一方面切割焦亡效应蛋白gasdermin D(GSDMD)形成GSDMD-N端,而GSDMD-N端在细胞膜上寡聚形成微孔,导致细胞渗透压的变化,进而发生肿胀直至细胞膜破裂;另一方面裂解白介素-1β(interleukin-1β,IL-1β)前体使其成为成熟的IL-1β,并通过GSDMD小孔释放从而引发过度炎症反应,造成组织损伤[6-7].细胞焦亡在BPD的发生和发展中起到的重要作用已经被许多研究证实.NOD样受体热蛋白结构域相关蛋白3(NOD-like receptor thermal protein domain associated protein 3,NLRP3)炎症小体及NLRP1炎症小体被认为在BPD的发生和发展中发挥着重要作用[8-9].此外,抑制caspase-1活性或IL-1β分泌均可改善BPD新生鼠的肺损伤[9-10].而作为肺发育主体的肺泡上皮细胞,其焦亡引发的呼吸系统疾病也被广泛报道[11-12].Necrosulfonamide(NSA)是一种GSDMD特异性抑制剂,不仅能抑制GSDMD蛋白的切割,还能抑制GSDMD-N端寡聚化和膜穿孔的形成[13-14].但抑制焦亡效应蛋白GSDMD是否可以抑制肺泡上皮细胞的焦亡从而改善BPD,目前尚不清楚.因此,本研究拟探究GSDMD抑制剂NSA对羊膜腔内注射脂多糖(lipopolysaccharide,LPS)诱导的BPD新生大鼠肺发育的影响以及对其肺泡上皮细胞焦亡的抑制作用. ...
... 细胞焦亡是一种炎性含半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,caspase)依赖性的细胞死亡,其特征是细胞肿胀、溶解和多种炎症因子的释放.多种炎症小体的激活可活化caspase.活性caspase一方面切割焦亡效应蛋白gasdermin D(GSDMD)形成GSDMD-N端,而GSDMD-N端在细胞膜上寡聚形成微孔,导致细胞渗透压的变化,进而发生肿胀直至细胞膜破裂;另一方面裂解白介素-1β(interleukin-1β,IL-1β)前体使其成为成熟的IL-1β,并通过GSDMD小孔释放从而引发过度炎症反应,造成组织损伤[6-7].细胞焦亡在BPD的发生和发展中起到的重要作用已经被许多研究证实.NOD样受体热蛋白结构域相关蛋白3(NOD-like receptor thermal protein domain associated protein 3,NLRP3)炎症小体及NLRP1炎症小体被认为在BPD的发生和发展中发挥着重要作用[8-9].此外,抑制caspase-1活性或IL-1β分泌均可改善BPD新生鼠的肺损伤[9-10].而作为肺发育主体的肺泡上皮细胞,其焦亡引发的呼吸系统疾病也被广泛报道[11-12].Necrosulfonamide(NSA)是一种GSDMD特异性抑制剂,不仅能抑制GSDMD蛋白的切割,还能抑制GSDMD-N端寡聚化和膜穿孔的形成[13-14].但抑制焦亡效应蛋白GSDMD是否可以抑制肺泡上皮细胞的焦亡从而改善BPD,目前尚不清楚.因此,本研究拟探究GSDMD抑制剂NSA对羊膜腔内注射脂多糖(lipopolysaccharide,LPS)诱导的BPD新生大鼠肺发育的影响以及对其肺泡上皮细胞焦亡的抑制作用. ...
1
... 细胞焦亡是一种炎性含半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,caspase)依赖性的细胞死亡,其特征是细胞肿胀、溶解和多种炎症因子的释放.多种炎症小体的激活可活化caspase.活性caspase一方面切割焦亡效应蛋白gasdermin D(GSDMD)形成GSDMD-N端,而GSDMD-N端在细胞膜上寡聚形成微孔,导致细胞渗透压的变化,进而发生肿胀直至细胞膜破裂;另一方面裂解白介素-1β(interleukin-1β,IL-1β)前体使其成为成熟的IL-1β,并通过GSDMD小孔释放从而引发过度炎症反应,造成组织损伤[6-7].细胞焦亡在BPD的发生和发展中起到的重要作用已经被许多研究证实.NOD样受体热蛋白结构域相关蛋白3(NOD-like receptor thermal protein domain associated protein 3,NLRP3)炎症小体及NLRP1炎症小体被认为在BPD的发生和发展中发挥着重要作用[8-9].此外,抑制caspase-1活性或IL-1β分泌均可改善BPD新生鼠的肺损伤[9-10].而作为肺发育主体的肺泡上皮细胞,其焦亡引发的呼吸系统疾病也被广泛报道[11-12].Necrosulfonamide(NSA)是一种GSDMD特异性抑制剂,不仅能抑制GSDMD蛋白的切割,还能抑制GSDMD-N端寡聚化和膜穿孔的形成[13-14].但抑制焦亡效应蛋白GSDMD是否可以抑制肺泡上皮细胞的焦亡从而改善BPD,目前尚不清楚.因此,本研究拟探究GSDMD抑制剂NSA对羊膜腔内注射脂多糖(lipopolysaccharide,LPS)诱导的BPD新生大鼠肺发育的影响以及对其肺泡上皮细胞焦亡的抑制作用. ...
2
... 细胞焦亡是一种炎性含半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,caspase)依赖性的细胞死亡,其特征是细胞肿胀、溶解和多种炎症因子的释放.多种炎症小体的激活可活化caspase.活性caspase一方面切割焦亡效应蛋白gasdermin D(GSDMD)形成GSDMD-N端,而GSDMD-N端在细胞膜上寡聚形成微孔,导致细胞渗透压的变化,进而发生肿胀直至细胞膜破裂;另一方面裂解白介素-1β(interleukin-1β,IL-1β)前体使其成为成熟的IL-1β,并通过GSDMD小孔释放从而引发过度炎症反应,造成组织损伤[6-7].细胞焦亡在BPD的发生和发展中起到的重要作用已经被许多研究证实.NOD样受体热蛋白结构域相关蛋白3(NOD-like receptor thermal protein domain associated protein 3,NLRP3)炎症小体及NLRP1炎症小体被认为在BPD的发生和发展中发挥着重要作用[8-9].此外,抑制caspase-1活性或IL-1β分泌均可改善BPD新生鼠的肺损伤[9-10].而作为肺发育主体的肺泡上皮细胞,其焦亡引发的呼吸系统疾病也被广泛报道[11-12].Necrosulfonamide(NSA)是一种GSDMD特异性抑制剂,不仅能抑制GSDMD蛋白的切割,还能抑制GSDMD-N端寡聚化和膜穿孔的形成[13-14].但抑制焦亡效应蛋白GSDMD是否可以抑制肺泡上皮细胞的焦亡从而改善BPD,目前尚不清楚.因此,本研究拟探究GSDMD抑制剂NSA对羊膜腔内注射脂多糖(lipopolysaccharide,LPS)诱导的BPD新生大鼠肺发育的影响以及对其肺泡上皮细胞焦亡的抑制作用. ...
... 细胞焦亡是一种炎性含半胱氨酸的天冬氨酸蛋白水解酶(cysteinyl aspartate specific proteinase,caspase)依赖性的细胞死亡,其特征是细胞肿胀、溶解和多种炎症因子的释放.多种炎症小体的激活可活化caspase.活性caspase一方面切割焦亡效应蛋白gasdermin D(GSDMD)形成GSDMD-N端,而GSDMD-N端在细胞膜上寡聚形成微孔,导致细胞渗透压的变化,进而发生肿胀直至细胞膜破裂;另一方面裂解白介素-1β(interleukin-1β,IL-1β)前体使其成为成熟的IL-1β,并通过GSDMD小孔释放从而引发过度炎症反应,造成组织损伤[6-7].细胞焦亡在BPD的发生和发展中起到的重要作用已经被许多研究证实.NOD样受体热蛋白结构域相关蛋白3(NOD-like receptor thermal protein domain associated protein 3,NLRP3)炎症小体及NLRP1炎症小体被认为在BPD的发生和发展中发挥着重要作用[8-9].此外,抑制caspase-1活性或IL-1β分泌均可改善BPD新生鼠的肺损伤[9-10].而作为肺发育主体的肺泡上皮细胞,其焦亡引发的呼吸系统疾病也被广泛报道[11-12].Necrosulfonamide(NSA)是一种GSDMD特异性抑制剂,不仅能抑制GSDMD蛋白的切割,还能抑制GSDMD-N端寡聚化和膜穿孔的形成[13-14].但抑制焦亡效应蛋白GSDMD是否可以抑制肺泡上皮细胞的焦亡从而改善BPD,目前尚不清楚.因此,本研究拟探究GSDMD抑制剂NSA对羊膜腔内注射脂多糖(lipopolysaccharide,LPS)诱导的BPD新生大鼠肺发育的影响以及对其肺泡上皮细胞焦亡的抑制作用. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}