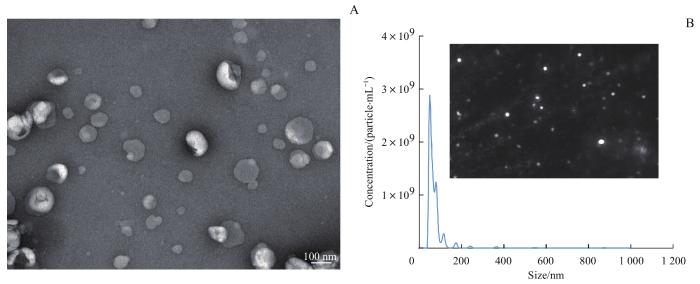
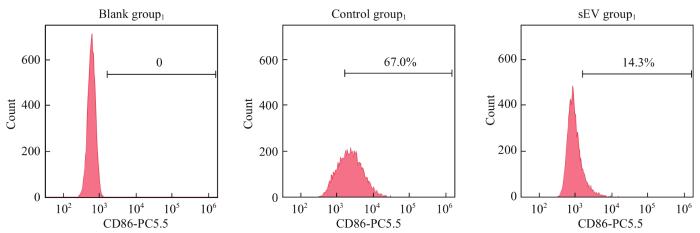
目的·探究人骨髓间充质干细胞(bone marrow mesenchymal stem cell,BMSC)来源的小细胞外囊泡(small extracellular vesicle,sEV)对小鼠破骨细胞分化和巨噬细胞极化的调控作用,以及对骨质疏松症小鼠的影响。方法·培养BMSC并通过差速离心法提取sEV,通过透射电子显微镜(transmission electron microscope,TEM)及纳米颗粒跟踪分析技术(nanoparticle tracking analysis,NTA)鉴定得到的sEV。通过巨噬细胞集落刺激因子(macrophage colony-stimulating factor,M-CSF)及核因子κB受体激活蛋白配体(receptor activator of nuclear factor-κB ligand,RANKL)刺激RAW264.7细胞以诱导形成破骨细胞,通过抗酒石酸酸性磷酸酶(tartrate-resistant acid phosphatase,TRAP)染色及鬼笔环肽染色检测sEV对破骨细胞分化的调控作用。通过荧光定量PCR检测sEV对破骨细胞标志基因环磷腺苷效应元件结合蛋白(cAMP-response element binding protein,CREB)、组织蛋白酶K(cathepsin K,CTSK)及c-Jun(Jun proto-oncogene)mRNA表达量的影响。使用脂多糖刺激RAW264.7细胞极化为M1型巨噬细胞;使用白细胞介素-4(interleukin-4,IL-4)及IL-13刺激RAW264.7细胞极化为M2型巨噬细胞。利用流式细胞术检测sEV对M1及M2型巨噬细胞极化的影响。通过微计算机断层扫描成像(micro-computed tomography,micro-CT)及TRAP染色观察sEV对骨质疏松症小鼠模型腰椎骨组织的影响。结果·TEM及NTA结果显示分离得到的sEV具有典型的球状结构,直径为30~150 nm。TRAP染色及鬼笔环肽染色结果显示,BMSC来源的sEV能够有效抑制RAW264.7细胞融合形成破骨细胞。PCR结果表明sEV能够降低CREB、CTSK和c-Jun mRNA的表达量(均P<0.05)。流式细胞术分析表明,BMSC来源的sEV能够抑制RAW264.7细胞极化为M1型巨噬细胞,促进其极化为M2型巨噬细胞。Micro-CT检测结果显示,sEV干预后模型小鼠腰椎骨小梁数量和骨体积分数显著高于未干预小鼠(均P<0.05);TRAP染色结果显示,sEV干预后腰椎组织中的破骨细胞数量减少。结论·人BMSC来源的sEV可以延缓骨质疏松小鼠的骨质流失,这可能与其抑制小鼠破骨细胞分化及促进M2型巨噬细胞极化的作用有关。
关键词:骨髓间充质干细胞
;
小细胞外囊泡
;
骨质疏松症
;
破骨细胞
;
巨噬细胞极化
Abstract
Objective ·To investigate the effects of small extracellular vesicles (sEVs) derived from human bone marrow mesenchymal stem cells (BMSCs) on the regulation of osteoclast differentiation and macrophage polarization in mice, and mouse model of osteoporosis. Methods ·BMSCs were cultured and sEVs were isolated through differential centrifugation. The isolated sEVs were identified by transmission electron microscopy (TEM) and nanoparticle tracking analysis (NTA). RAW264.7 cells were cultured and stimulated with macrophage colony-stimulating factor (M-CSF) and receptor activator of nuclear factor-κB ligand (RANKL) to differentiate the cells into osteoclasts. Tartrate-resistant acid phosphatase (TRAP) staining and phalloidin staining were performed to assess the effect of sEVs on osteoclast formation. The expression levels of osteoclast marker genes, i.e., cAMP-response element binding protein (CREB), cathepsin K (CTSK), and Jun proto-oncogene (c-Jun) were examined by real-time quantitative PCR. To polarize RAW264.7 cells to M1 phenotype, they were cultured with lipopolysaccharides; to polarize them to M2 phenotype, they were cultured with interleukin-4 (IL-4) and IL-13. Flow cytometry was performed to detect the effect of sEVs on macrophage polarization. Micro-computed tomography (micro-CT) and TRAP staining were performed to investigate the effect of sEVs on the bone tissues of lumbar vertebrae in osteoporosis mouse models. Results ·TEM and NTA demonstrated that the isolated sEVs had a typical globular structure with a diameter ranging from 30‒150 nm. TRAP staining and phalloidin staining showed that BMSC-derived sEVs inhibited the fusion of RAW264.7 cells to form osteoblasts. PCR revealed that sEVs could decrease the expression of CREB, CTSK, and c-Jun (all P<0.05). Flow cytometry analysis indicated that BMSC-derived sEVs inhibited RAW264.7 macrophages polarization to M1 phenotype and induced RAW264.7 macrophages polarization to M2 phenotype. Micro-CT indicated that the number of trabeculae and the bone volume fraction of lumbar vertebrae were significantly higher in the sEV-intervened group than those in the control group (both P<0.05). TRAP staining revealed a reduction of osteoclast number in the lumbar vertebrae after intervention with sEVs. Conclusion ·The sEVs from human BMSCs can delay bone loss in osteoporosis mice, which may be related to its effects of inhibiting osteoclast differentiation and promoting the polarization of M2 type macrophages.
LI Xuran, TAO Shicong, GUO Shangchun. Ameliorative effects on osteoporosis of small extracellular vesicles derived from bone marrow mesenchymal stem cells. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(4): 406-416 doi:10.3969/j.issn.1674-8115.2023.04.002
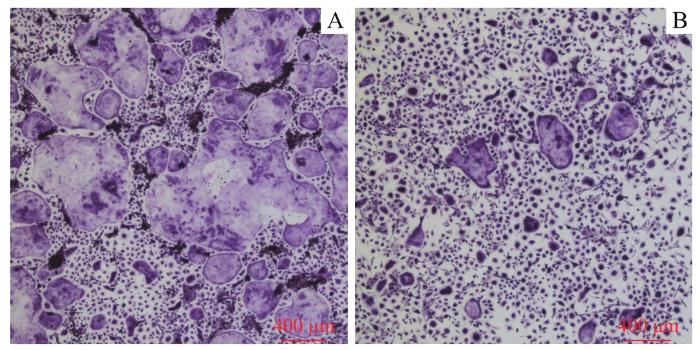
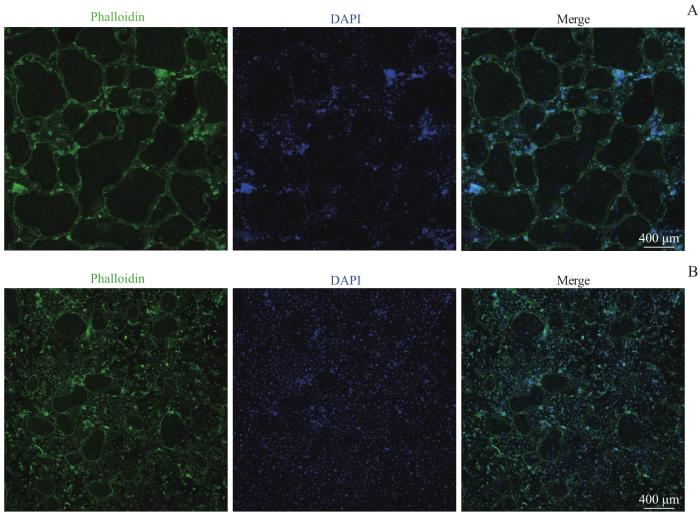
Note: A/B. The RAW264.7 cells were seeded at a high density (1.5×106 per well, A) and a low density (7.5×105 per well, B) and the formation of osteoclasts was observed at different time-points (×40). C/D. The results of TRAP staining of the cells seeded at a high density (C) and a low density (D) (×40).
Fig 2
Formation of osteoclasts and the results of TRAP staining
Note: A. The live imaging of the spines (left: the control group; right: the sEV group). B. The live imaging of the femurs and the tibias (left: the control group; right: the sEV group).
Fig 8
Observation of sEVs distribution in mouse bones by living imaging
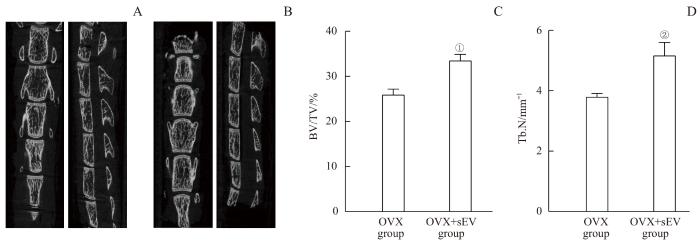
Note: A. The lumbar spine of the OVX group. B. The lumbar spine of the OVX+sEV group. C. The BV/TV values of 2 groups. D. The Tb.N values of 2 groups. ①P=0.002, ②P=0.005, compared with the OVX group.
Fig 9
Micro-CT imaging analysis and bone parameters of the osteoporosis mice and the sEV-intervened mice
GUO Shangchun and TAO Shicong were responsible for experimental design and paper revising. LI Xuran was responsible for experiments, data analyses, and paper writing. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
CHEATHAM S W, HANNEY W, KOLBER M, et al. Osteoporosis: exercise programming insight for the sports medicine professional[J]. Strength Cond J, 2017, 39: 2-13.
SI L, WINZENBERG T M, JIANG Q, et al. Projection of osteoporosis-related fractures and costs in China: 2010‒2050[J]. Osteoporos Int, 2015, 26(7): 1929-1937.
LEE C W, LIN H C, WANG B Y, et al. Ginkgolide B monotherapy reverses osteoporosis by regulating oxidative stress-mediated bone homeostasis[J]. Free Radic Biol Med, 2021, 168: 234-246.
LIU P, LEE S, KNOLL J, et al. Loss of menin in osteoblast lineage affects osteocyte-osteoclast crosstalk causing osteoporosis[J]. Cell Death Differ, 2017, 24(4): 672-682.
LEE K M, LEE C Y, ZHANG G, et al. Methylglyoxal activates osteoclasts through JNK pathway leading to osteoporosis[J]. Chem Biol Interact, 2019, 308: 147-154.
JACOME-GALARZA C E, PERCIN G I, MULLER J T, et al. Developmental origin, functional maintenance and genetic rescue of osteoclasts[J]. Nature, 2019, 568(7753): 541-545.
PESCE VIGLIETTI A I, GIAMBARTOLOMEI G H, DELPINO M V. Endocrine modulation of Brucella abortus-infected osteocytes function and osteoclastogenesis via modulation of RANKL/OPG[J]. Microbes Infect, 2019, 21(7): 287-295.
QIAN J, HE Y, ZHAO J, et al. IL4/IL4R signaling promotes the osteolysis in metastatic bone of CRC through regulating the proliferation of osteoclast precursors[J]. Mol Med, 2021, 27(1): 152.
CAI F Y, LIU S L, LEI Y X, et al. Epigallocatechin-3 gallate regulates macrophage subtypes and immunometabolism to ameliorate experimental autoimmune encephalomyelitis[J]. Cell Immunol, 2021, 368: 104421.
ZHANG Z G, ZHANG C Y, ZHANG S R. Irisin activates M1 macrophage and suppresses Th2-type immune response in rats with pelvic inflammatory disease[J]. Evid Based Complement Alternat Med, 2022, 2022: 5215915.
EOM J, YOO J, KIM J J, et al. Viperin deficiency promotes polarization of macrophages and secretion of M1 and M2 cytokines[J]. Immune Netw, 2018, 18(4): e32.
ZHANG W J, GUAN N, ZHANG X M, et al. Study on the imbalance of M1/M2 macrophage polarization in severe chronic periodontitis[J]. Technol Health Care, 2023, 31(1): 117-124.
WANG W H, LIU H, LIU T, et al. Insights into the role of macrophage polarization in the pathogenesis of osteoporosis[J]. Oxid Med Cell Longev, 2022, 2022: 2485959.
YU L, HU M, CUI X, et al. M1 macrophage-derived exosomes aggravate bone loss in postmenopausal osteoporosis via a microRNA-98/DUSP1/JNK axis[J]. Cell Biol Int, 2021, 45(12): 2452-2463.
LU Y P, LIU S S, YANG P P, et al. Exendin-4 and eldecalcitol synergistically promote osteogenic differentiation of bone marrow mesenchymal stem cells through M2 macrophages polarization via PI3K/AKT pathway[J]. Stem Cell Res Ther, 2022, 13(1): 113.
CHEN M, LIN W M, YE R, et al. PPARβ/δ agonist alleviates diabetic osteoporosis via regulating M1/M2 macrophage polarization[J]. Front Cell Dev Biol, 2021, 9: 753194.
LI M D, JIA J, LI S S, et al. Exosomes derived from tendon stem cells promote cell proliferation and migration through the TGF β signal pathway[J]. Biochem Biophys Res Commun, 2021, 536: 88-94.
YANG S D, GUO S, TONG S, et al. Promoting osteogenic differentiation of human adipose-derived stem cells by altering the expression of exosomal miRNA[J]. Stem Cells Int, 2019, 2019: 1351860.
ZHANG B B, ZHAXI D W, LI C, et al. M2 macrophagy-derived exosomal miRNA-26a-5p induces osteogenic differentiation of bone mesenchymal stem cells[J]. J Orthop Surg Res, 2022, 17(1): 137.
ZHU G C, CHEN W, TANG C Y, et al. Knockout and double knockout of cathepsin K and Mmp9 reveals a novel function of cathepsin K as a regulator of osteoclast gene expression and bone homeostasis[J]. Int J Biol Sci, 2022, 18(14): 5522-5538.
HE F T, LUO S H, LIU S J, et al. Zanthoxylum bungeanum seed oil inhibits RANKL-induced osteoclastogenesis by suppressing ERK/c-JUN/NFATc1 pathway and regulating cell cycle arrest in RAW264.7 cells[J]. J Ethnopharmacol, 2022, 289: 115094.
KUMAR A, HUGHES T M, CRAFT S, et al. A novel approach to isolate brain-cell-derived exosomes from plasma to better understand pathogenesis of Alzheimer's disease[J]. Alzheimer's Dement, 2020, 16(Suppl 4): e044894.
LI K, WONG D K, HONG K Y, et al. Cushioned-density gradient ultracentrifugation (C-DGUC): a refined and high performance method for the isolation, characterization, and use of exosomes[J]. Methods Mol Biol, 2018, 1740: 69-83.
HELWA I, CAI J W, DREWRY M D, et al. A comparative study of serum exosome isolation using differential ultracentrifugation and three commercial reagents[J]. PLoS One, 2017, 12(1): e0170628.
DING M, WANG C, LU X L, et al. Comparison of commercial exosome isolation kits for circulating exosomal microRNA profiling[J]. Anal Bioanal Chem, 2018, 410(16): 3805-3814.
LI K, XIU C M, ZHOU Q, et al. A dual role of cholesterol in osteogenic differentiation of bone marrow stromal cells[J]. J Cell Physiol, 2019, 234(3): 2058-2066.
CHE Y T, YANG J Z, TANG F, et al. New function of cholesterol oxidation products involved in osteoporosis pathogenesis[J]. Int J Mol Sci, 2022, 23(4): 2020.
LI K Q, CHEN S H, CAI P Y, et al. MiRNA-483-5p is involved in the pathogenesis of osteoporosis by promoting osteoclast differentiation[J]. Mol Cell Probes, 2020, 49: 101479.
PARK E, LEE C G, LIM E, et al. Osteoprotective effects of loganic acid on osteoblastic and osteoclastic cells and osteoporosis-induced mice[J]. Int J Mol Sci, 2020, 22(1): 233.
LAI G H, ZHAO R L, ZHUANG W D, et al. BMSC-derived exosomal miR-27a-3p and miR-196b-5p regulate bone remodeling in ovariectomized rats[J]. PeerJ, 2022, 10: e13744.
SONG H Y, LI X Q, ZHAO Z C, et al. Reversal of osteoporotic activity by endothelial cell-secreted bone targeting and biocompatible exosomes[J]. Nano Lett, 2019, 19(5): 3040-3048.
CHEN X T, WAN Z, YANG L, et al. Exosomes derived from reparative M2-like macrophages prevent bone loss in murine periodontitis models via IL-10 mRNA[J]. J Nanobiotechnology, 2022, 20(1): 110.
ZHU L F, LI L, WANG X Q, et al. M1 macrophages regulate TLR4/AP1 via paracrine to promote alveolar bone destruction in periodontitis[J]. Oral Dis, 2019, 25(8): 1972-1982.
LIANG B L, WANG H C, WU D, et al. Macrophage M1/M2 polarization dynamically adapts to changes in microenvironment and modulates alveolar bone remodeling after dental implantation[J]. J Leukoc Biol, 2021, 110(3): 433-447.
SHI M S, WANG C, WANG Y L, et al. Deproteinized bovine bone matrix induces osteoblast differentiation via macrophage polarization[J]. J Biomed Mater Res A, 2018, 106(5): 1236-1246.
SHI C, YUAN F, LI Z L, et al. MSN@IL-4 sustainingly mediates macrophagocyte M2 polarization and relieves osteoblast damage via NF-κB pathway-associated apoptosis[J]. Biomed Res Int, 2022, 2022: 2898729.
Horibe K, Hara M, Nakamura H. M2-like macrophage infiltration and transforming growth factor-β secretion during socket healing process in mice[J]. Arch Oral Biol, 2021, 123: 105042.
WANG X Y, JI Q B, HU W H, et al. Isobavachalcone prevents osteoporosis by suppressing activation of ERK and NF-κB pathways and M1 polarization of macrophages[J]. Int Immunopharmacol, 2021, 94: 107370.
LI Z K, ZHU X D, XU R J, et al. Deacylcynaropicrin inhibits RANKL-induced osteoclastogenesis by inhibiting NF-κB and MAPK and promoting M2 polarization of macrophages[J]. Front Pharmacol, 2019, 10: 599.
YAO M Y, CUI B, ZHANG W H, et al. Exosomal miR-21 secreted by IL-1β-primed-mesenchymal stem cells induces macrophage M2 polarization and ameliorates sepsis[J]. Life Sci, 2021, 264: 118658.
LI R, LI D Z, WANG H N, et al. Exosomes from adipose-derived stem cells regulate M1/M2 macrophage phenotypic polarization to promote bone healing via miR-451a/MIF[J]. Stem Cell Res Ther, 2022, 13(1): 149.
LI R, ZHAO K C, RUAN Q, et al. Bone marrow mesenchymal stem cell-derived exosomal microRNA-124-3p attenuates neurological damage in spinal cord ischemia-reperfusion injury by downregulating Ern1 and promoting M2 macrophage polarization[J]. Arthritis Res Ther, 2020, 22(1): 75.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}