Ovarian cancer is a malignant tumor of the female reproductive system with the highest mortality, and involves the aberrant regulation of multiple molecular signaling pathways. As a highly conserved molecular pathway, Wnt signaling pathway plays an important role in embryonic development, tissue homeostasis, and tumorigenesis. Wnt signaling pathway includes canonical Wnt/β-catenin pathway and non-canonical pathway, and the latter mainly includes Wnt/planar cell polarity (Wnt/PCP) pathway and Wnt/Ca2+ pathway. Previous studies have mainly focused on the relationship between the canonical Wnt pathway and tumor progression. Recently, the non-canonical Wnt pathway has gradually received attention, and related researches have enriched the understanding of the non-canonical Wnt pathway in physiological and pathological processes such as tissue development and tumorigenesis. The existing studies suggest that the nonclassical Wnt pathway is abnormally regulated in ovarian cancer and is closely related to the staging and prognosis of ovarian cancer. Non-classical Wnt pathway plays an important role in many biological processes such as proliferation, migration and invasion of ovarian cancer cells, and the changes of this pathway are also related to chemotherapy resistance of ovarian cancer. This article reviews the role of the non-canonical Wnt pathway in ovarian cancer, and discusses the research progress of targeted therapy based on the non-canonical Wnt pathway, aiming to provide new ideas for the development of novel targeted drugs.
Keywords:ovarian cancer
;
non-canonical Wnt pathway
;
cancer progression
;
chemotherapy resistance
;
targeted therapy
ZHOU Wanzhen, TENG Yincheng. Research progress of the role of non-canonical Wnt signaling pathway in ovarian cancer and its potential therapeutic implications. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(8): 1056-1063 doi:10.3969/j.issn.1674-8115.2023.08.015
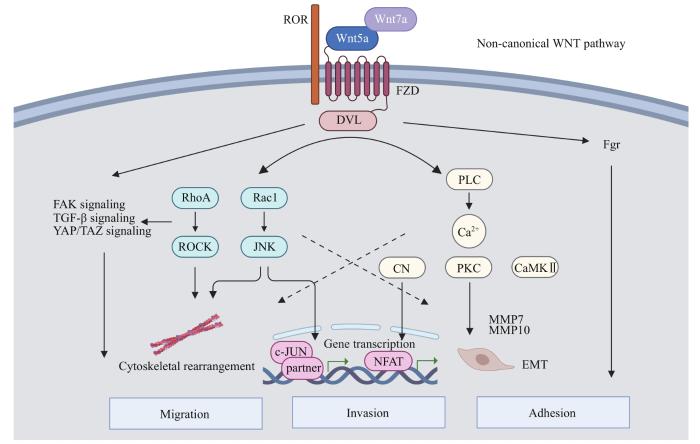
非经典信号通路主要包括2种机制——Wnt/PCP通路和Wnt/Ca2+通路,通常由非经典Wnt蛋白激活。Wnt/PCP通路参与原肠胚形成、神经前体细胞分化以及内耳毛细胞定位等多种生物学过程,通常被认为与细胞极性、排列和迁移相关[5-6]。在Wnt/PCP通路中,Wnt与FZD、ROR的结合可活化DVL,并可激活形态发生紊乱蛋白相关激活因子1(dishevelled-associated activator of morphogenesis 1,DAAM1),诱导Ras同系物家族成员A(Ras homolog family member A,RhoA)及其下游Rho相关卷曲螺旋蛋白激酶(Rho-associated coiled-coil forming protein kinase,ROCK)的磷酸化,从而实现细胞肌动蛋白骨架重排;DVL还可通过直接激活Ras相关的C3肉毒素底物1(Ras-related C3 botulinum toxin substrate 1,Rac1)诱导肌动蛋白聚合及伪足形成,并活化c-Jun N末端蛋白激酶(c-Jun N-terminal protein kinase,JNK),促进c-Jun磷酸化并与Fos及ATF等蛋白家族成员二聚化构成激活蛋白1(activator protein 1,AP-1)转录复合物,进而启动MMP-2、MMP-9、尿激酶型纤溶酶原激活剂(urokinase type plasminogen activator,uPA)、CCND2等靶基因转录[7]。Wnt/Ca2+通路的调控及功能作用较为复杂且研究相对有限。现有文献资料[8]表明其在调节细胞骨架重排、细胞黏附以及胚胎背-腹轴模式建立等过程中可发挥重要作用。Wnt-FZD复合物可通过激活磷脂酶C(phospholipase C,PLC)提高细胞内Ca2+浓度,进一步激活钙调神经磷酸酶(calcineurin,CN)及多种钙依赖性激酶,如钙/钙调蛋白依赖的蛋白激酶Ⅱ(calcium/calmodulin-dependent protein kinase Ⅱ,CaMKⅡ)和蛋白激酶C(protein kinase C,PKC)[4]。CN可诱导活化的T细胞核内因子(nuclear factor of activated T cell,NFAT)入核,启动靶基因转录。
Wnt/PCP通路可通过激活ROCK及Rac1诱导肌动蛋白骨架重塑,参与细胞形态和细胞运动的调节。RhoA-ROCK以及Rac1-JNK级联介导的丝状伪足及板状伪足形成是Wnt5a诱导肿瘤细胞迁移的关键过程[5,30]。在体外模型中,Wnt5a重组蛋白刺激卵巢癌细胞迁移及侵袭能力增强,并观察到经Wnt5a处理的卵巢癌细胞丝状伪足及板状伪足数目及长度增加的显著细胞形态学改变[26]。Dickkopf相关蛋白4(Dickkopf-related protein 4,DDK4)是经典Wnt通路的抑制因子,但可激活非经典的Wnt/PCP通路,在Wnt经典和非经典通路的调控中发挥重要作用。研究[31]表明,DKK4的mRNA和蛋白水平在卵巢癌中均显著增高,利用siRNA敲低卵巢癌细胞株中的DKK4可通过抑制c-Jun、JNK的磷酸化和肌动蛋白丝的形成来抑制细胞迁移。
Fig 1
Non-canonical Wnt signaling pathway regulates ovarian cancer cell migration and invasion
5 非经典Wnt通路与卵巢癌细胞化疗耐药
经典Wnt通路在卵巢癌铂耐药中的研究已取得了显著进展,而非经典Wnt通路在卵巢癌耐药中的作用也日益受到关注。VESKIMÄE等[34]研究发现,与铂敏感卵巢癌患者相比,Wnt5a、ROR1和ROR2的蛋白表达水平在铂耐药患者的肿瘤组织中显著增高。在卵巢癌细胞中过表达Wnt5a可显著降低细胞对紫杉醇、奥沙利铂、5-氟尿嘧啶、表柔比星和依托泊苷的化学敏感性,而敲低Wnt5a则可增加对上述药物的化学敏感性[10]。HUNG等[35]发现,Wnt5a在肿瘤多重耐药细胞中高表达并可通过激活蛋白激酶A来上调ATP结合盒亚家族B成员1(ATP binding cassette subfamily B member 1,ABCB1)的表达。ABCB1可以ATP依赖的形式将药物泵出细胞,降低胞内化疗药物的浓度,介导多药耐药。非经典Wnt通路的其他组分对卵巢癌化疗耐药也起到重要作用。ROR1和ROR2在A2780顺铂耐药株(A2780/cisR)中的表达显著高于亲本株,特异性敲低ROR1和ROR2可抑制卵巢癌铂耐药细胞的迁移和侵袭,并增强细胞对顺铂的敏感性[12]。ZHANG等[36]发现,DVL1在卵巢癌紫杉醇耐药细胞株(A2780/Taxol)中高表达,抑制DVL1活性可增加紫杉醇诱导的细胞毒性。因此,Wnt5a及下游关键蛋白的表达水平可能是卵巢癌化疗反应的潜在生物标志物。
TENG Yincheng was responsible for the guidance, formulation and review of the paper framework. ZHOU Wanzhen was responsible for literature reading and paper writing. Both authors have read the last version of paper and consented for submission.
利益冲突声明
2位作者声明不存在利益冲突。
COMPETING INTERESTS
Both authors disclose no relevant conflict of interests.
SUNG H, FERLAY J, SIEGEL R L, et al. Global cancer statistics 2020: globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries[J]. CA Cancer J Clin, 2021, 71(3): 209-249.
NGUYEN V H L, HOUGH R, BERNAUDO S, et al. Wnt/β-catenin signalling in ovarian cancer: insights into its hyperactivation and function in tumorigenesis[J]. J Ovarian Res, 2019, 12(1): 122.
VANDERVORST K, HATAKEYAMA J, BERG A, et al. Cellular and molecular mechanisms underlying planar cell polarity pathway contributions to cancer malignancy[J]. Semin Cell Dev Biol, 2018, 81: 78-87.
FORD C E, PUNNIA-MOORTHY G, HENRY C E, et al. The non-canonical Wnt ligand, Wnt5a, is upregulated and associated with epithelial to mesenchymal transition in epithelial ovarian cancer[J]. Gynecol Oncol, 2014, 134(2): 338-345.
PENG C J, ZHANG X L, YU H L, et al. Wnt5a as a predictor in poor clinical outcome of patients and a mediator in chemoresistance of ovarian cancer[J]. Int J Gynecol Cancer, 2011, 21(2): 280-288.
JIN P, SONG Y, YU G Y. The role of abnormal methylation of Wnt5a gene promoter regions in human epithelial ovarian cancer: a clinical and experimental study[J]. Anal Cell Pathol (Amst), 2018, 2018: 6567081.
HENRY C E, LLAMOSAS E, DJORDJEVIC A, et al. Migration and invasion is inhibited by silencing ROR1 and ROR2 in chemoresistant ovarian cancer[J]. Oncogenesis, 2016, 5(5): e226.
KARIN-KUJUNDZIC V, KARDUM V, SOLA I M, et al. Dishevelled family proteins in serous ovarian carcinomas: a clinicopathologic and molecular study[J]. APMIS, 2020, 128(3): 201-210.
LIU R, CHENG J L, CHEN Y N, et al. Potential role and prognostic importance of dishevelled-2 in epithelial ovarian cancer[J]. Int J Gynaecol Obstet, 2017, 138(3): 304-310.
XU W W, GU J J, REN Q L, et al. NFATC1 promotes cell growth and tumorigenesis in ovarian cancer up-regulating c-Myc through ERK1/2/p38 MAPK signal pathway[J]. Tumor Biol, 2016, 37(4): 4493-4500.
XIN B, JI K Q, LIU Y S, et al. NFAT overexpression correlates with CA72-4 and poor prognosis of ovarian clear-cell carcinoma subtype[J]. Reprod Sci, 2021, 28(3): 745-756.
CHEN S, WANG J, GOU W F, et al. The involvement of RhoA and Wnt5a in the tumorigenesis and progression of ovarian epithelial carcinoma[J]. Int J Mol Sci, 2013, 14(12): 24187-24199.
CHEHOVER M, REICH R, DAVIDSON B. Expression of Wnt pathway molecules is associated with disease outcome in metastatic high-grade serous carcinoma[J]. Virchows Arch, 2020, 477(2): 249-258.
AZIMIAN-ZAVAREH V, DEHGHANI-GHOBADI Z, EBRAHIMI M, et al. Wnt5a modulates integrin expression in a receptor-dependent manner in ovarian cancer cells[J]. Sci Rep, 2021, 11(1): 5885.
HENRY C E, EMMANUEL C, LAMBIE N, et al. Distinct patterns of stromal and tumor expression of ROR1 and ROR2 in histological subtypes of epithelial ovarian cancer[J]. Transl Oncol, 2017, 10(3): 346-356.
ZHANG S P, CUI B, LAI H, et al. Ovarian cancer stem cells express ROR1, which can be targeted for anti-cancer-stem-cell therapy[J]. Proc Natl Acad Sci USA, 2014, 111(48): 17266-17271.
FANG X, CHEN C Q, XIA F Z, et al. CD274 promotes cell cycle entry of leukemia-initiating cells through JNK/cyclin D2 signaling[J]. J Hematol Oncol, 2016, 9(1): 124.
ASEM M, YOUNG A M, OYAMA C, et al. Host Wnt5a potentiates microenvironmental regulation of ovarian cancer metastasis[J]. Cancer Res, 2020, 80(5): 1156-1170.
LUO M, ZHOU L, ZHAN S J, et al. ALPL regulates the aggressive potential of high grade serous ovarian cancer cells via a non-canonical Wnt pathway[J]. Biochem Biophys Res Commun, 2019, 513(2): 528-533.
QI H, SUN B C, ZHAO X L, et al. Wnt5a promotes vasculogenic mimicry and epithelial-mesenchymal transition via protein kinase Cα in epithelial ovarian cancer[J]. Oncol Rep, 2014, 32(2): 771-779.
AL-ALEM L F, MCCORD L A, SOUTHARD R C, et al. Activation of the PKC pathway stimulates ovarian cancer cell proliferation, migration, and expression of MMP7 and MMP10[J]. Biol Reprod, 2013, 89(3): 73.
WANG S Z, WEI H, ZHANG S L. Dickkopf-4 is frequently overexpressed in epithelial ovarian carcinoma and promotes tumor invasion[J]. BMC Cancer, 2017, 17(1): 455.
DEHGHANI-GHOBADI Z, SHEIKH HASANI S, AREFIAN E, et al. Wnt5a and TGFβ1 converges through YAP1 activity and integrin alpha v up-regulation promoting epithelial to mesenchymal transition in ovarian cancer cells and mesothelial cell activation[J]. Cells, 2022, 11(2): 237.
VESKIMÄE K, SCARAVILLI M, NIININEN W, et al. Expression analysis of platinum sensitive and resistant epithelial ovarian cancer patient samples reveals new candidates for targeted therapies[J]. Transl Oncol, 2018, 11(5): 1160-1170.
HUNG T H, HSU S C, CHENG C Y, et al. Wnt5a regulates ABCB1 expression in multidrug-resistant cancer cells through activation of the non-canonical PKA/β-catenin pathway[J]. Oncotarget, 2014, 5(23): 12273-12290.
ZHANG K, SONG H X, YANG P, et al. Silencing dishevelled-1 sensitizes paclitaxel-resistant human ovarian cancer cells via AKT/GSK-3β/β-catenin signalling[J]. Cell Prolif, 2015, 48(2): 249-258.
HUANG L, JIN Y, FENG S J, et al. Role of Wnt/β-catenin, Wnt/c-Jun N-terminal kinase and Wnt/Ca2+ pathways in cisplatin-induced chemoresistance in ovarian cancer[J]. Exp Ther Med, 2016, 12(6): 3851-3858.
HUANG W, YANG S, CHENG Y S, et al. Terfenadine resensitizes doxorubicin activity in drug-resistant ovarian cancer cells via an inhibition of CaMKⅡ/CREB1 mediated ABCB1 expression[J]. Front Oncol, 2022, 12: 1068443.
JENEI V, SHERWOOD V, HOWLIN J, et al. A t-butyloxycarbonyl-modified Wnt5a-derived hexapeptide functions as a potent antagonist of Wnt5a-dependent melanoma cell invasion[J]. Proc Natl Acad Sci USA, 2009, 106(46): 19473-19478.
MOORE K N, GUNDERSON C C, SABBATINI P, et al. A phase 1b dose escalation study of ipafricept (OMP54F28) in combination with paclitaxel and carboplatin in patients with recurrent platinum-sensitive ovarian cancer[J]. Gynecol Oncol, 2019, 154(2): 294-301.
CHOI M Y, WIDHOPF G F Ⅱ, WU C C, et al. Pre-clinical specificity and safety of UC-961, a first-in-class monoclonal antibody targeting ROR1[J]. Clin Lymphoma Myeloma Leuk, 2015, 15: S167-S169.
CHOI M Y, WIDHOPF G F Ⅱ, GHIA E M, et al. Phase Ⅰ trial: cirmtuzumab inhibits ROR1 signaling and stemness signatures in patients with chronic lymphocytic leukemia[J]. Cell Stem Cell, 2018, 22(6): 951-959.e3.
GHIA E M, RASSENTI L Z, CHOI M Y, et al. High expression level of ROR1 and ROR1-signaling associates with venetoclax resistance in chronic lymphocytic leukemia[J]. Leukemia, 2022, 36(6): 1609-1618.
ZHANG S P, ZHANG H, GHIA E M, et al. Inhibition of chemotherapy resistant breast cancer stem cells by a ROR1 specific antibody[J]. Proc Natl Acad Sci USA, 2019, 116(4): 1370-1377.
LIU D L, KAUFMANN G F, BREITMEYER J B, et al. The anti-ROR1 monoclonal antibody zilovertamab inhibits the proliferation of ovarian and endometrial cancer cells[J]. Pharmaceutics, 2022, 14(4): 837.
WU D, YU X Y, WANG J, et al. Ovarian cancer stem cells with high ROR1 expression serve as a new prophylactic vaccine for ovarian cancer[J]. J Immunol Res, 2019, 2019: 9394615.
SRIVASTAVA S, FURLAN S N, JAEGER-RUCKSTUHL C A, et al. Immunogenic chemotherapy enhances recruitment of CAR-T cells to lung tumors and improves antitumor efficacy when combined with checkpoint blockade[J]. Cancer Cell, 2021, 39(2): 193-208.e10.
JOSHI N, LIU D L, DICKSON K A, et al. An organotypic model of high-grade serous ovarian cancer to test the anti-metastatic potential of ROR2 targeted Polyion complex nanoparticles[J]. J Mater Chem B, 2021, 9(44): 9123-9135.
CHEN X Y, CHEN Y M, LIN X Y, et al. The drug combination of SB202190 and SP600125 significantly inhibit the growth and metastasis of olaparib-resistant ovarian cancer cell[J]. Curr Pharm Biotechnol, 2018, 19(6): 506-513.
OHTA T, TAKAHASHI T, SHIBUYA T, et al. Inhibition of the Rho/ROCK pathway enhances the efficacy of cisplatin through the blockage of hypoxia-inducible factor-1α in human ovarian cancer cells[J]. Cancer Biol Ther, 2012, 13(1): 25-33.
... 非经典信号通路主要包括2种机制——Wnt/PCP通路和Wnt/Ca2+通路,通常由非经典Wnt蛋白激活.Wnt/PCP通路参与原肠胚形成、神经前体细胞分化以及内耳毛细胞定位等多种生物学过程,通常被认为与细胞极性、排列和迁移相关[5-6].在Wnt/PCP通路中,Wnt与FZD、ROR的结合可活化DVL,并可激活形态发生紊乱蛋白相关激活因子1(dishevelled-associated activator of morphogenesis 1,DAAM1),诱导Ras同系物家族成员A(Ras homolog family member A,RhoA)及其下游Rho相关卷曲螺旋蛋白激酶(Rho-associated coiled-coil forming protein kinase,ROCK)的磷酸化,从而实现细胞肌动蛋白骨架重排;DVL还可通过直接激活Ras相关的C3肉毒素底物1(Ras-related C3 botulinum toxin substrate 1,Rac1)诱导肌动蛋白聚合及伪足形成,并活化c-Jun N末端蛋白激酶(c-Jun N-terminal protein kinase,JNK),促进c-Jun磷酸化并与Fos及ATF等蛋白家族成员二聚化构成激活蛋白1(activator protein 1,AP-1)转录复合物,进而启动MMP-2、MMP-9、尿激酶型纤溶酶原激活剂(urokinase type plasminogen activator,uPA)、CCND2等靶基因转录[7].Wnt/Ca2+通路的调控及功能作用较为复杂且研究相对有限.现有文献资料[8]表明其在调节细胞骨架重排、细胞黏附以及胚胎背-腹轴模式建立等过程中可发挥重要作用.Wnt-FZD复合物可通过激活磷脂酶C(phospholipase C,PLC)提高细胞内Ca2+浓度,进一步激活钙调神经磷酸酶(calcineurin,CN)及多种钙依赖性激酶,如钙/钙调蛋白依赖的蛋白激酶Ⅱ(calcium/calmodulin-dependent protein kinase Ⅱ,CaMKⅡ)和蛋白激酶C(protein kinase C,PKC)[4].CN可诱导活化的T细胞核内因子(nuclear factor of activated T cell,NFAT)入核,启动靶基因转录. ...
2
... 非经典信号通路主要包括2种机制——Wnt/PCP通路和Wnt/Ca2+通路,通常由非经典Wnt蛋白激活.Wnt/PCP通路参与原肠胚形成、神经前体细胞分化以及内耳毛细胞定位等多种生物学过程,通常被认为与细胞极性、排列和迁移相关[5-6].在Wnt/PCP通路中,Wnt与FZD、ROR的结合可活化DVL,并可激活形态发生紊乱蛋白相关激活因子1(dishevelled-associated activator of morphogenesis 1,DAAM1),诱导Ras同系物家族成员A(Ras homolog family member A,RhoA)及其下游Rho相关卷曲螺旋蛋白激酶(Rho-associated coiled-coil forming protein kinase,ROCK)的磷酸化,从而实现细胞肌动蛋白骨架重排;DVL还可通过直接激活Ras相关的C3肉毒素底物1(Ras-related C3 botulinum toxin substrate 1,Rac1)诱导肌动蛋白聚合及伪足形成,并活化c-Jun N末端蛋白激酶(c-Jun N-terminal protein kinase,JNK),促进c-Jun磷酸化并与Fos及ATF等蛋白家族成员二聚化构成激活蛋白1(activator protein 1,AP-1)转录复合物,进而启动MMP-2、MMP-9、尿激酶型纤溶酶原激活剂(urokinase type plasminogen activator,uPA)、CCND2等靶基因转录[7].Wnt/Ca2+通路的调控及功能作用较为复杂且研究相对有限.现有文献资料[8]表明其在调节细胞骨架重排、细胞黏附以及胚胎背-腹轴模式建立等过程中可发挥重要作用.Wnt-FZD复合物可通过激活磷脂酶C(phospholipase C,PLC)提高细胞内Ca2+浓度,进一步激活钙调神经磷酸酶(calcineurin,CN)及多种钙依赖性激酶,如钙/钙调蛋白依赖的蛋白激酶Ⅱ(calcium/calmodulin-dependent protein kinase Ⅱ,CaMKⅡ)和蛋白激酶C(protein kinase C,PKC)[4].CN可诱导活化的T细胞核内因子(nuclear factor of activated T cell,NFAT)入核,启动靶基因转录. ...
... Wnt/PCP通路可通过激活ROCK及Rac1诱导肌动蛋白骨架重塑,参与细胞形态和细胞运动的调节.RhoA-ROCK以及Rac1-JNK级联介导的丝状伪足及板状伪足形成是Wnt5a诱导肿瘤细胞迁移的关键过程[5,30].在体外模型中,Wnt5a重组蛋白刺激卵巢癌细胞迁移及侵袭能力增强,并观察到经Wnt5a处理的卵巢癌细胞丝状伪足及板状伪足数目及长度增加的显著细胞形态学改变[26].Dickkopf相关蛋白4(Dickkopf-related protein 4,DDK4)是经典Wnt通路的抑制因子,但可激活非经典的Wnt/PCP通路,在Wnt经典和非经典通路的调控中发挥重要作用.研究[31]表明,DKK4的mRNA和蛋白水平在卵巢癌中均显著增高,利用siRNA敲低卵巢癌细胞株中的DKK4可通过抑制c-Jun、JNK的磷酸化和肌动蛋白丝的形成来抑制细胞迁移. ...
1
... 非经典信号通路主要包括2种机制——Wnt/PCP通路和Wnt/Ca2+通路,通常由非经典Wnt蛋白激活.Wnt/PCP通路参与原肠胚形成、神经前体细胞分化以及内耳毛细胞定位等多种生物学过程,通常被认为与细胞极性、排列和迁移相关[5-6].在Wnt/PCP通路中,Wnt与FZD、ROR的结合可活化DVL,并可激活形态发生紊乱蛋白相关激活因子1(dishevelled-associated activator of morphogenesis 1,DAAM1),诱导Ras同系物家族成员A(Ras homolog family member A,RhoA)及其下游Rho相关卷曲螺旋蛋白激酶(Rho-associated coiled-coil forming protein kinase,ROCK)的磷酸化,从而实现细胞肌动蛋白骨架重排;DVL还可通过直接激活Ras相关的C3肉毒素底物1(Ras-related C3 botulinum toxin substrate 1,Rac1)诱导肌动蛋白聚合及伪足形成,并活化c-Jun N末端蛋白激酶(c-Jun N-terminal protein kinase,JNK),促进c-Jun磷酸化并与Fos及ATF等蛋白家族成员二聚化构成激活蛋白1(activator protein 1,AP-1)转录复合物,进而启动MMP-2、MMP-9、尿激酶型纤溶酶原激活剂(urokinase type plasminogen activator,uPA)、CCND2等靶基因转录[7].Wnt/Ca2+通路的调控及功能作用较为复杂且研究相对有限.现有文献资料[8]表明其在调节细胞骨架重排、细胞黏附以及胚胎背-腹轴模式建立等过程中可发挥重要作用.Wnt-FZD复合物可通过激活磷脂酶C(phospholipase C,PLC)提高细胞内Ca2+浓度,进一步激活钙调神经磷酸酶(calcineurin,CN)及多种钙依赖性激酶,如钙/钙调蛋白依赖的蛋白激酶Ⅱ(calcium/calmodulin-dependent protein kinase Ⅱ,CaMKⅡ)和蛋白激酶C(protein kinase C,PKC)[4].CN可诱导活化的T细胞核内因子(nuclear factor of activated T cell,NFAT)入核,启动靶基因转录. ...
1
... 非经典信号通路主要包括2种机制——Wnt/PCP通路和Wnt/Ca2+通路,通常由非经典Wnt蛋白激活.Wnt/PCP通路参与原肠胚形成、神经前体细胞分化以及内耳毛细胞定位等多种生物学过程,通常被认为与细胞极性、排列和迁移相关[5-6].在Wnt/PCP通路中,Wnt与FZD、ROR的结合可活化DVL,并可激活形态发生紊乱蛋白相关激活因子1(dishevelled-associated activator of morphogenesis 1,DAAM1),诱导Ras同系物家族成员A(Ras homolog family member A,RhoA)及其下游Rho相关卷曲螺旋蛋白激酶(Rho-associated coiled-coil forming protein kinase,ROCK)的磷酸化,从而实现细胞肌动蛋白骨架重排;DVL还可通过直接激活Ras相关的C3肉毒素底物1(Ras-related C3 botulinum toxin substrate 1,Rac1)诱导肌动蛋白聚合及伪足形成,并活化c-Jun N末端蛋白激酶(c-Jun N-terminal protein kinase,JNK),促进c-Jun磷酸化并与Fos及ATF等蛋白家族成员二聚化构成激活蛋白1(activator protein 1,AP-1)转录复合物,进而启动MMP-2、MMP-9、尿激酶型纤溶酶原激活剂(urokinase type plasminogen activator,uPA)、CCND2等靶基因转录[7].Wnt/Ca2+通路的调控及功能作用较为复杂且研究相对有限.现有文献资料[8]表明其在调节细胞骨架重排、细胞黏附以及胚胎背-腹轴模式建立等过程中可发挥重要作用.Wnt-FZD复合物可通过激活磷脂酶C(phospholipase C,PLC)提高细胞内Ca2+浓度,进一步激活钙调神经磷酸酶(calcineurin,CN)及多种钙依赖性激酶,如钙/钙调蛋白依赖的蛋白激酶Ⅱ(calcium/calmodulin-dependent protein kinase Ⅱ,CaMKⅡ)和蛋白激酶C(protein kinase C,PKC)[4].CN可诱导活化的T细胞核内因子(nuclear factor of activated T cell,NFAT)入核,启动靶基因转录. ...
1
... 非经典信号通路主要包括2种机制——Wnt/PCP通路和Wnt/Ca2+通路,通常由非经典Wnt蛋白激活.Wnt/PCP通路参与原肠胚形成、神经前体细胞分化以及内耳毛细胞定位等多种生物学过程,通常被认为与细胞极性、排列和迁移相关[5-6].在Wnt/PCP通路中,Wnt与FZD、ROR的结合可活化DVL,并可激活形态发生紊乱蛋白相关激活因子1(dishevelled-associated activator of morphogenesis 1,DAAM1),诱导Ras同系物家族成员A(Ras homolog family member A,RhoA)及其下游Rho相关卷曲螺旋蛋白激酶(Rho-associated coiled-coil forming protein kinase,ROCK)的磷酸化,从而实现细胞肌动蛋白骨架重排;DVL还可通过直接激活Ras相关的C3肉毒素底物1(Ras-related C3 botulinum toxin substrate 1,Rac1)诱导肌动蛋白聚合及伪足形成,并活化c-Jun N末端蛋白激酶(c-Jun N-terminal protein kinase,JNK),促进c-Jun磷酸化并与Fos及ATF等蛋白家族成员二聚化构成激活蛋白1(activator protein 1,AP-1)转录复合物,进而启动MMP-2、MMP-9、尿激酶型纤溶酶原激活剂(urokinase type plasminogen activator,uPA)、CCND2等靶基因转录[7].Wnt/Ca2+通路的调控及功能作用较为复杂且研究相对有限.现有文献资料[8]表明其在调节细胞骨架重排、细胞黏附以及胚胎背-腹轴模式建立等过程中可发挥重要作用.Wnt-FZD复合物可通过激活磷脂酶C(phospholipase C,PLC)提高细胞内Ca2+浓度,进一步激活钙调神经磷酸酶(calcineurin,CN)及多种钙依赖性激酶,如钙/钙调蛋白依赖的蛋白激酶Ⅱ(calcium/calmodulin-dependent protein kinase Ⅱ,CaMKⅡ)和蛋白激酶C(protein kinase C,PKC)[4].CN可诱导活化的T细胞核内因子(nuclear factor of activated T cell,NFAT)入核,启动靶基因转录. ...
... 经典Wnt通路在卵巢癌铂耐药中的研究已取得了显著进展,而非经典Wnt通路在卵巢癌耐药中的作用也日益受到关注.VESKIMÄE等[34]研究发现,与铂敏感卵巢癌患者相比,Wnt5a、ROR1和ROR2的蛋白表达水平在铂耐药患者的肿瘤组织中显著增高.在卵巢癌细胞中过表达Wnt5a可显著降低细胞对紫杉醇、奥沙利铂、5-氟尿嘧啶、表柔比星和依托泊苷的化学敏感性,而敲低Wnt5a则可增加对上述药物的化学敏感性[10].HUNG等[35]发现,Wnt5a在肿瘤多重耐药细胞中高表达并可通过激活蛋白激酶A来上调ATP结合盒亚家族B成员1(ATP binding cassette subfamily B member 1,ABCB1)的表达.ABCB1可以ATP依赖的形式将药物泵出细胞,降低胞内化疗药物的浓度,介导多药耐药.非经典Wnt通路的其他组分对卵巢癌化疗耐药也起到重要作用.ROR1和ROR2在A2780顺铂耐药株(A2780/cisR)中的表达显著高于亲本株,特异性敲低ROR1和ROR2可抑制卵巢癌铂耐药细胞的迁移和侵袭,并增强细胞对顺铂的敏感性[12].ZHANG等[36]发现,DVL1在卵巢癌紫杉醇耐药细胞株(A2780/Taxol)中高表达,抑制DVL1活性可增加紫杉醇诱导的细胞毒性.因此,Wnt5a及下游关键蛋白的表达水平可能是卵巢癌化疗反应的潜在生物标志物. ...
... 经典Wnt通路在卵巢癌铂耐药中的研究已取得了显著进展,而非经典Wnt通路在卵巢癌耐药中的作用也日益受到关注.VESKIMÄE等[34]研究发现,与铂敏感卵巢癌患者相比,Wnt5a、ROR1和ROR2的蛋白表达水平在铂耐药患者的肿瘤组织中显著增高.在卵巢癌细胞中过表达Wnt5a可显著降低细胞对紫杉醇、奥沙利铂、5-氟尿嘧啶、表柔比星和依托泊苷的化学敏感性,而敲低Wnt5a则可增加对上述药物的化学敏感性[10].HUNG等[35]发现,Wnt5a在肿瘤多重耐药细胞中高表达并可通过激活蛋白激酶A来上调ATP结合盒亚家族B成员1(ATP binding cassette subfamily B member 1,ABCB1)的表达.ABCB1可以ATP依赖的形式将药物泵出细胞,降低胞内化疗药物的浓度,介导多药耐药.非经典Wnt通路的其他组分对卵巢癌化疗耐药也起到重要作用.ROR1和ROR2在A2780顺铂耐药株(A2780/cisR)中的表达显著高于亲本株,特异性敲低ROR1和ROR2可抑制卵巢癌铂耐药细胞的迁移和侵袭,并增强细胞对顺铂的敏感性[12].ZHANG等[36]发现,DVL1在卵巢癌紫杉醇耐药细胞株(A2780/Taxol)中高表达,抑制DVL1活性可增加紫杉醇诱导的细胞毒性.因此,Wnt5a及下游关键蛋白的表达水平可能是卵巢癌化疗反应的潜在生物标志物. ...
... Wnt/PCP通路可通过激活ROCK及Rac1诱导肌动蛋白骨架重塑,参与细胞形态和细胞运动的调节.RhoA-ROCK以及Rac1-JNK级联介导的丝状伪足及板状伪足形成是Wnt5a诱导肿瘤细胞迁移的关键过程[5,30].在体外模型中,Wnt5a重组蛋白刺激卵巢癌细胞迁移及侵袭能力增强,并观察到经Wnt5a处理的卵巢癌细胞丝状伪足及板状伪足数目及长度增加的显著细胞形态学改变[26].Dickkopf相关蛋白4(Dickkopf-related protein 4,DDK4)是经典Wnt通路的抑制因子,但可激活非经典的Wnt/PCP通路,在Wnt经典和非经典通路的调控中发挥重要作用.研究[31]表明,DKK4的mRNA和蛋白水平在卵巢癌中均显著增高,利用siRNA敲低卵巢癌细胞株中的DKK4可通过抑制c-Jun、JNK的磷酸化和肌动蛋白丝的形成来抑制细胞迁移. ...
... Wnt/PCP通路可通过激活ROCK及Rac1诱导肌动蛋白骨架重塑,参与细胞形态和细胞运动的调节.RhoA-ROCK以及Rac1-JNK级联介导的丝状伪足及板状伪足形成是Wnt5a诱导肿瘤细胞迁移的关键过程[5,30].在体外模型中,Wnt5a重组蛋白刺激卵巢癌细胞迁移及侵袭能力增强,并观察到经Wnt5a处理的卵巢癌细胞丝状伪足及板状伪足数目及长度增加的显著细胞形态学改变[26].Dickkopf相关蛋白4(Dickkopf-related protein 4,DDK4)是经典Wnt通路的抑制因子,但可激活非经典的Wnt/PCP通路,在Wnt经典和非经典通路的调控中发挥重要作用.研究[31]表明,DKK4的mRNA和蛋白水平在卵巢癌中均显著增高,利用siRNA敲低卵巢癌细胞株中的DKK4可通过抑制c-Jun、JNK的磷酸化和肌动蛋白丝的形成来抑制细胞迁移. ...
1
... Wnt/PCP通路可通过激活ROCK及Rac1诱导肌动蛋白骨架重塑,参与细胞形态和细胞运动的调节.RhoA-ROCK以及Rac1-JNK级联介导的丝状伪足及板状伪足形成是Wnt5a诱导肿瘤细胞迁移的关键过程[5,30].在体外模型中,Wnt5a重组蛋白刺激卵巢癌细胞迁移及侵袭能力增强,并观察到经Wnt5a处理的卵巢癌细胞丝状伪足及板状伪足数目及长度增加的显著细胞形态学改变[26].Dickkopf相关蛋白4(Dickkopf-related protein 4,DDK4)是经典Wnt通路的抑制因子,但可激活非经典的Wnt/PCP通路,在Wnt经典和非经典通路的调控中发挥重要作用.研究[31]表明,DKK4的mRNA和蛋白水平在卵巢癌中均显著增高,利用siRNA敲低卵巢癌细胞株中的DKK4可通过抑制c-Jun、JNK的磷酸化和肌动蛋白丝的形成来抑制细胞迁移. ...
... 经典Wnt通路在卵巢癌铂耐药中的研究已取得了显著进展,而非经典Wnt通路在卵巢癌耐药中的作用也日益受到关注.VESKIMÄE等[34]研究发现,与铂敏感卵巢癌患者相比,Wnt5a、ROR1和ROR2的蛋白表达水平在铂耐药患者的肿瘤组织中显著增高.在卵巢癌细胞中过表达Wnt5a可显著降低细胞对紫杉醇、奥沙利铂、5-氟尿嘧啶、表柔比星和依托泊苷的化学敏感性,而敲低Wnt5a则可增加对上述药物的化学敏感性[10].HUNG等[35]发现,Wnt5a在肿瘤多重耐药细胞中高表达并可通过激活蛋白激酶A来上调ATP结合盒亚家族B成员1(ATP binding cassette subfamily B member 1,ABCB1)的表达.ABCB1可以ATP依赖的形式将药物泵出细胞,降低胞内化疗药物的浓度,介导多药耐药.非经典Wnt通路的其他组分对卵巢癌化疗耐药也起到重要作用.ROR1和ROR2在A2780顺铂耐药株(A2780/cisR)中的表达显著高于亲本株,特异性敲低ROR1和ROR2可抑制卵巢癌铂耐药细胞的迁移和侵袭,并增强细胞对顺铂的敏感性[12].ZHANG等[36]发现,DVL1在卵巢癌紫杉醇耐药细胞株(A2780/Taxol)中高表达,抑制DVL1活性可增加紫杉醇诱导的细胞毒性.因此,Wnt5a及下游关键蛋白的表达水平可能是卵巢癌化疗反应的潜在生物标志物. ...
1
... 经典Wnt通路在卵巢癌铂耐药中的研究已取得了显著进展,而非经典Wnt通路在卵巢癌耐药中的作用也日益受到关注.VESKIMÄE等[34]研究发现,与铂敏感卵巢癌患者相比,Wnt5a、ROR1和ROR2的蛋白表达水平在铂耐药患者的肿瘤组织中显著增高.在卵巢癌细胞中过表达Wnt5a可显著降低细胞对紫杉醇、奥沙利铂、5-氟尿嘧啶、表柔比星和依托泊苷的化学敏感性,而敲低Wnt5a则可增加对上述药物的化学敏感性[10].HUNG等[35]发现,Wnt5a在肿瘤多重耐药细胞中高表达并可通过激活蛋白激酶A来上调ATP结合盒亚家族B成员1(ATP binding cassette subfamily B member 1,ABCB1)的表达.ABCB1可以ATP依赖的形式将药物泵出细胞,降低胞内化疗药物的浓度,介导多药耐药.非经典Wnt通路的其他组分对卵巢癌化疗耐药也起到重要作用.ROR1和ROR2在A2780顺铂耐药株(A2780/cisR)中的表达显著高于亲本株,特异性敲低ROR1和ROR2可抑制卵巢癌铂耐药细胞的迁移和侵袭,并增强细胞对顺铂的敏感性[12].ZHANG等[36]发现,DVL1在卵巢癌紫杉醇耐药细胞株(A2780/Taxol)中高表达,抑制DVL1活性可增加紫杉醇诱导的细胞毒性.因此,Wnt5a及下游关键蛋白的表达水平可能是卵巢癌化疗反应的潜在生物标志物. ...
1
... 经典Wnt通路在卵巢癌铂耐药中的研究已取得了显著进展,而非经典Wnt通路在卵巢癌耐药中的作用也日益受到关注.VESKIMÄE等[34]研究发现,与铂敏感卵巢癌患者相比,Wnt5a、ROR1和ROR2的蛋白表达水平在铂耐药患者的肿瘤组织中显著增高.在卵巢癌细胞中过表达Wnt5a可显著降低细胞对紫杉醇、奥沙利铂、5-氟尿嘧啶、表柔比星和依托泊苷的化学敏感性,而敲低Wnt5a则可增加对上述药物的化学敏感性[10].HUNG等[35]发现,Wnt5a在肿瘤多重耐药细胞中高表达并可通过激活蛋白激酶A来上调ATP结合盒亚家族B成员1(ATP binding cassette subfamily B member 1,ABCB1)的表达.ABCB1可以ATP依赖的形式将药物泵出细胞,降低胞内化疗药物的浓度,介导多药耐药.非经典Wnt通路的其他组分对卵巢癌化疗耐药也起到重要作用.ROR1和ROR2在A2780顺铂耐药株(A2780/cisR)中的表达显著高于亲本株,特异性敲低ROR1和ROR2可抑制卵巢癌铂耐药细胞的迁移和侵袭,并增强细胞对顺铂的敏感性[12].ZHANG等[36]发现,DVL1在卵巢癌紫杉醇耐药细胞株(A2780/Taxol)中高表达,抑制DVL1活性可增加紫杉醇诱导的细胞毒性.因此,Wnt5a及下游关键蛋白的表达水平可能是卵巢癌化疗反应的潜在生物标志物. ...



{kind=link}
{kind=link}