An integrated prognostic model of nuclear-encoded mitochondrial gene signature and clinical information for hepatocellular carcinoma
Aishanjiang Kedeerya,1,2, FU Yi2, LAI Donglin2,3, WU Hailong,2,4, GONG Wei,1,5
1.Department of General Surgery, Xinhua Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai 200092, China
2.Collaborative Innovation Center for Biomedicine, Shanghai University of Medicine and Health Sciences, Shanghai 201318, China
3.The First Affiliated Hospital, Jiangxi Medical College, Nanchang University, Nanchang 330006, China
4.School of Pharmacy, Shanghai University of Medicine and Health Sciences, Shanghai 201318, China
5.Shanghai Key Laboratory of Biliary Tract Disease Research, Research Institute of Biliary Tract Disease, Shanghai Jiao Tong University School of Medicine, Shanghai Research Center of Biliary Tract Disease, Shanghai 200092, China
Objective ·To establish a prognostic model for the overall survival (OS) of hepatocellular carcinoma (HCC) based on mitochondrial genes and clinical information. Methods ·The gene expression and the clinical data of 369 HCC patients and 50 controls with normal liver were downloaded from The Cancer Genome Atlas (TCGA) database. The nuclear-encoded mitochondrial genes (NEMGs) were obtained from the MitoCarta3.0 database. The "DESeq2" R package and univariate Cox analysis were used to select NEMGs [ubiquinol cytochrome C reductase hinge protein (UQCRH),ATP citrate lyase (ACLY),phosphoenolpyruvate carboxykinase 2 (PCK2), Bcl-2 homologous antagonist/killer1 (BAK1), Bcl-2-associated X protein (BAX) andBcl-2/adenovirus E1B interacting protein 3-like (BNIP3L)] in HCC that were associated with OS of HCC and participated in dysregulation of oxidative phosphorylation, tricarboxylic acid cycle and cell apoptosis. Multivariate Cox analysis was applied to select independent risk factors for OS of HCC. A comprehensive prognostic model and a prognostic nomogram with 6-NEMG risk characteristics and TNM staging were established. By using the median of prognostic scores as a cut-off, HCC patients were classified into low-risk and high-risk group. Kaplan-Meier survival curve analysis was conducted and log-rank test was performed to evaluate the survival rates between the low-risk and high-risk group. The area under the curve (AUC) values of receiver operating characteristic (ROC) curve were calculated via using the "timeROC" package. The prognostic model for HCC was validated by using the GEO HCC cohort (GSE14520) for 1, 3 and 5 years. Finally, the relative expression level of 6-NEMG was validated in 34 clinical samples of HCC from Xinhua Hospital, Shanghai Jiao Tong University School of Medicine by using real-time quantitative polymerase chain reaction (qPCR) method. Results ·Compared to 6-NEMG risk signature only (AUCs for 1, 3 and 5 years were 0.77, 0.66 and 0.65, respectively) or TNM stage only (AUCs for 1, 3 and 5 years were 0.66, 0.67 and 0.63, respectively), ROC curve analysis showed that this integrated prognostic model displayed better predictive performance for 1-year (AUC, 0.78), 3-year (AUC, 0.73) and 5-year (AUC, 0.69) OS of HCC. The Kaplan-Meier survival curve analysis showed that the OS of HCC patients in the high-risk group was significantly worse than that in the low-risk group (P=0.001). In addition, predictive performance of the prognostic model (AUC for 1, 3 and 5 years is 0.67, 0.66 and 0.74, respectively) and prognostic differences between the high-risk and low-risk group (P=0.001) were further validated in GEO (GSE14520) external cohort, and these results were consistent with the TCGA data. In addition to BNIP3L, dysregulation of five other NEMGs in the clinical HCC cohort was validated. The correlation analysis in GSE14520 and HCC clinical cohort showed a positive correlation between prognosis score and the size and number of tumors. Conclusion ·A new prognostic model that combines 6-NEMG risk characteristics with TNM staging for predicting OS in HCC patients was constructed and validated. This model may help improve the prognosis prediction of HCC patients.
Aishanjiang Kedeerya, FU Yi, LAI Donglin, WU Hailong, GONG Wei. An integrated prognostic model of nuclear-encoded mitochondrial gene signature and clinical information for hepatocellular carcinoma. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2024, 44(1): 1-12 doi:10.3969/j.issn.1674-8115.2024.01.001
在全球范围内,原发性肝癌是最具侵袭性和难以治疗的恶性肿瘤之一[1]。肝细胞癌(hepatocellular carcinoma,HCC)约占所有原发性肝癌病例的90%[2]。尽管早期和晚期HCC的治疗方法都有所改善[3-4],但近几十年来,许多国家的发病率和死亡率仍然很高[5]。由于预后不良,HCC目前是全球第四大常见的癌症相关死亡原因,5年生存率低于21%[6]。因此,建立一个能够准确预测HCC预后的模型对于改善HCC预后至关重要。虽然许多分类系统,如美国癌症联合委员会(American Joint Committee on Cancer,AJCC)的TNM分期(American Joint Committee on Cancer-Tumor Node Metastasis Staging)系统、巴塞罗那临床肝癌(Barcelona Clinic Liver Cancer,BCLC)分期和意大利肝癌评分(Cancer of the Liver Italian Program,CLIP)系统,已广泛应用于临床评价HCC患者的预后,但这些分类系统主要是基于临床病理特征,没有考虑分子标志物的重要预后作用[7]。
对包括年龄、性别、甲胎蛋白(α-fetoprotein,AFP)和TNM分期等重要的临床病理特征以及deNEMG的风险特征通过“forestplot”和“survival”包进行单因素和多因素Cox回归分析,获得独立的预后因素。使用“rms”和“nomogramEx”软件包建立了一个基于独立预后因素的预后模型和预后列线图。根据该预后模型中涉及的独立预后因素的系数,制定公式来计算每个患者的预后评分。以预后评分的中位数作为临界值,将HCC患者分为高风险组和低风险组。采用“survival”和“survminer”包进行Kaplan-Meier生存曲线分析,并采用log-rank检验评估低风险组和高风险组之间的OS。使用“timeROC”软件包计算受试者操作特征(receiver operating characteristic,ROC)曲线下面积(area under the curve,AUC)。
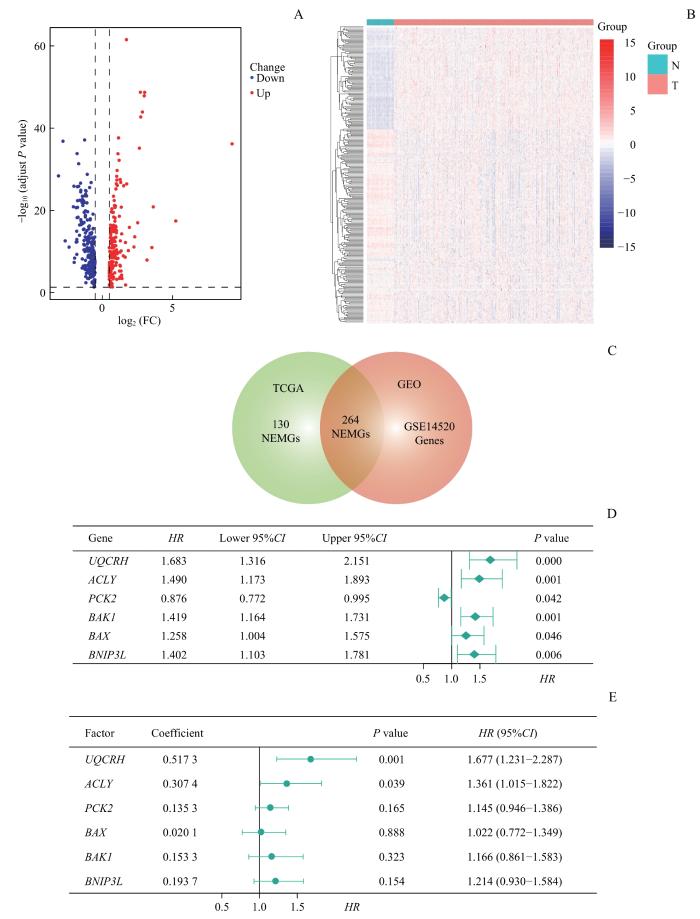
Note: A. DEGs between HCC and adjacent paired normal tissues. Red dots represent significant upregulation and blue dots represent significant downregulation of DEGs in HCC tissues. B. Heatmap of 264 deNEMGs in TCGA cohort. N—adjacent paired normal tissues; T—HCC tumor tissues. C. Venn diagram showing differently expressed deNEMGs between tumor and adjacent tissues. D. Univariate Cox regression analysis of 6 significant prognostic NEMGs associated with OXPHOS, TCA cycle and cell apoptosis. E. Multivariate Cox analysis of 6-NEMG risk signature.
Fig 1
Identification of prognosis-related NEMGs and construction of a 6-NEMG-based risk signature
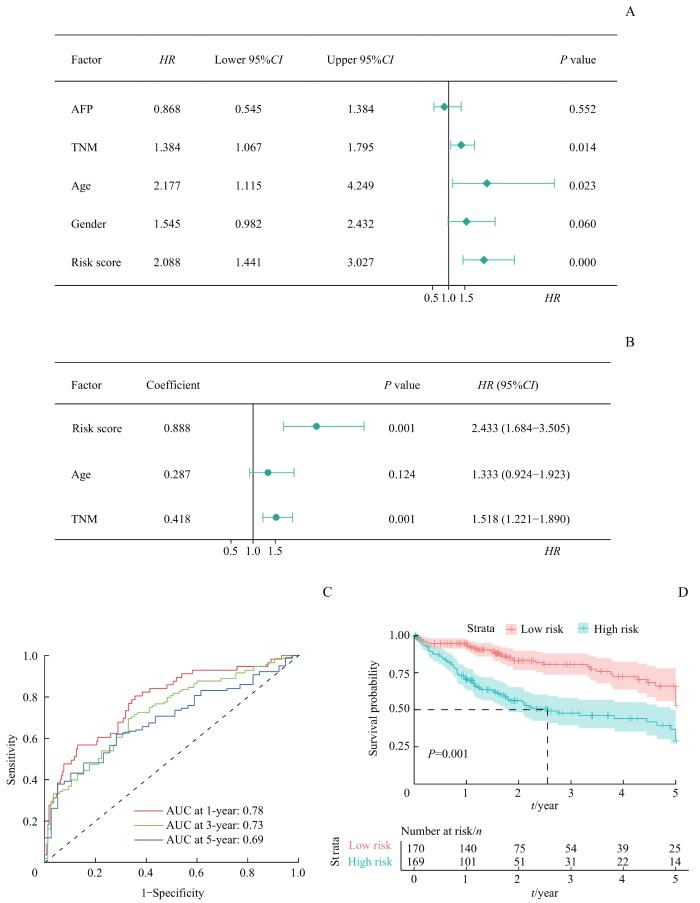
Note: A/B. Univariate (A) and multivariate (B) Cox analysis of risk score and clinicopathological features for OS of HCC. C. ROC analysis for the prognostic model at 1-, 3- and 5-year survival in the TCGA-LIHC cohort. D. Kaplan-Meier survival analysis of OS in the high- and low-risk group in the TCGA-LIHC cohort.
Fig 2
Construction of a prognostic model integrating the 6-NEMG risk signature with TNM stage
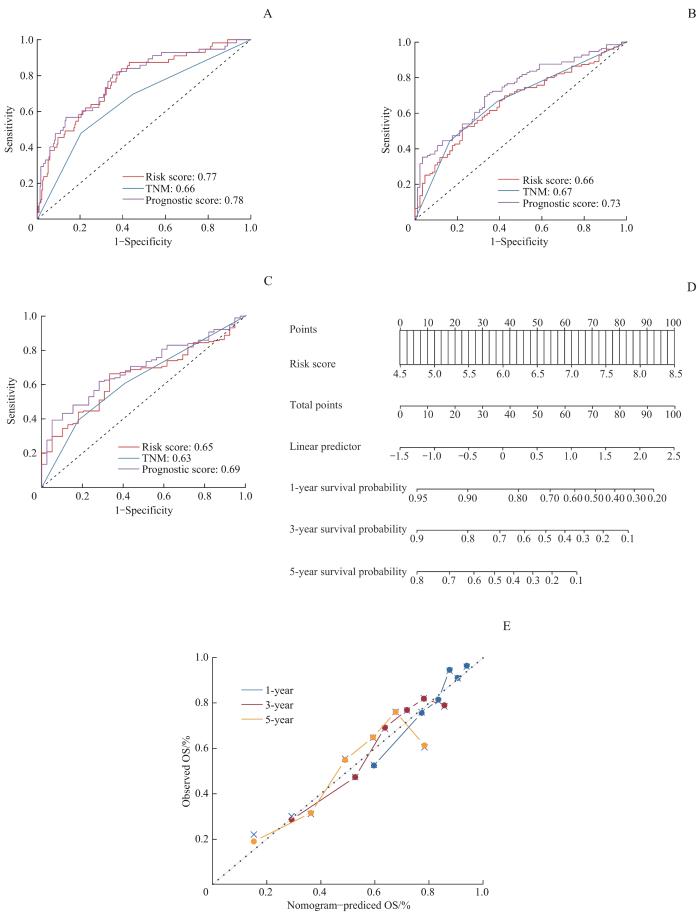
Note: A‒C. ROC analysis of the prognostic score, risk score and TNM stage for 1-year OS (A), 3-year OS (B) and 5-year OS (C). D. Nomogram constructed by the prognostic score to predict OS of HCC. E. Calibration curves of the prognostic score-derived nomogram for 1-year, 3-year and 5-year OS of HCC.
Fig 3
Establishment of the prognostic score-derived nomogram based on TCGA cohort
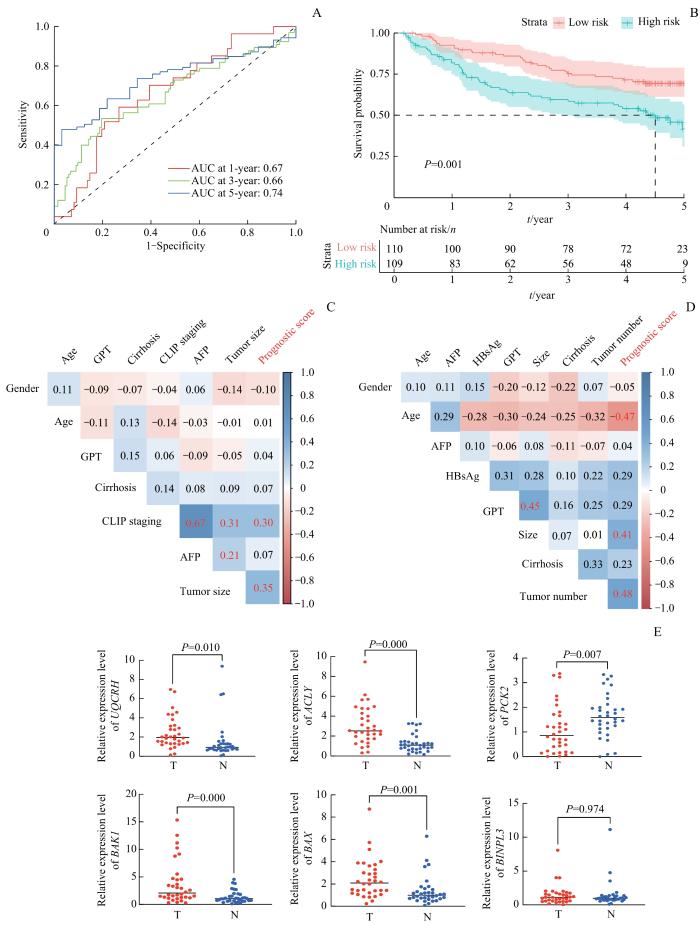
Note: A. ROC analysis of the prognostic model for OS of HCC in GSE14520. B. Kaplan-Meier analysis of OS in HCC patients of GSE14520 which were stratified into high- and low-risk groups by this prognostic model. C. Correlation analysis between clinicopathological features and prognostic score in HCC patients of GSE14520. The correlation coefficient in red indicates P<0.05. D. Correlation analysis between clinicopathological features and prognostic score in 34-pair HCC and matched normal tissues. The correlation coefficient in red indicates P<0.05. E. Gene expression levels of the 6 NEMGs involved in this prognostic model in 34-pair HCC tumor tissues (T) and matched normal tissues (N).
Fig 4
Validation of the prognostic model in GEO and clinical HCC cohort
本研究着重研究OXPHOS、TCA循环和细胞凋亡这3个通路中的预后相关基因,对95个与HCC预后相关的deNEMG和参与3种有丝分裂通路的NEMG取交集,发现仅6个参与3种有丝分裂通路的deNEMG(UQCRH、ACLY、PCK2、BAK1、BAX和BNIP3L)与HCC的预后密切相关。因此本研究利用这6个基因来构建基于NEMG的风险模型。在这6个NEMG中,UQCRH编码一个定位于线粒体和细胞核的铰链蛋白。UQCRH作为泛素-细胞色素C还原酶复合物(也称为OXPHOS通路的复合物Ⅲ)的主要亚基之一,负责复合物Ⅲ的细胞色素C和C1之间的电子转移。事实上,在肺腺癌[27]和HCC[28]中已经有关于UQCRH表达上调的报道。此外,UQCRH的表达与肿瘤大小、分化程度和有无血管侵袭相关,并与HCC患者的不良预后密切相关[28]。ACLY和PCK2参与TCA循环,其中ACLY形成一种细胞质同源四聚体酶,催化TCA循环中的中间体柠檬酸盐转化为草酰乙酸和乙酰辅酶A[29]。由于乙酰辅酶A是新脂肪生成和胆固醇生物合成的重要底物之一,因此ACLY是一种重要的脂肪生成酶,在结直肠癌、非小细胞肺癌、乳腺癌、膀胱癌和HCC等癌症类型中高表达[30-32]。同时,在HCC患者中,ACLY的高表达与不良预后、肿瘤干性和转移相关[33]。PCK2编码一种线粒体磷酸烯醇式丙酮酸羧激酶,该酶催化TCA循环衍生的草酰乙酸转化为磷酸烯醇式丙酮酸,并伴随GTP水解。PCK2在某些癌症类型,如肺癌、前列腺癌、乳腺癌、宫颈癌和睾丸癌中经常上调和激活[34-35],可能以此对抗肿瘤微环境中的代谢饥饿状态[36]。此外,既往研究[37]表明,PCK2的下调与HCC的预后和患癌时间有关,并且PCK2的异位过表达可能通过诱导HCC细胞的葡萄糖缺乏而降低细胞活力。BAX、BAK1和BNIP3L是凋亡相关基因,其中BAX和BAK1是线粒体外膜通透性的决定性关键因素[38-40]。BNIP3L是线粒体吞噬的双重调节因子,而线粒体吞噬是清除受损线粒体的过程[41]。据报道BNIP3L与BAX和BAK相互作用,导致线粒体外膜通透性增强和凋亡加快[42]。同时,BNIP3L介导的线粒体吞噬通过糖酵解代谢重编程,促进乙型肝炎病毒X蛋白(hepatitis B virus X protein,HBx)诱导的HCC的肿瘤干性[42]。
All experimental protocols in this study were reviewed and approved by Ethics Committee of Xinhua Hospital, Shanghai Jiao Tong University School of Medicine (Approval Letter No. XHEC-D-2023-021). Consent letters have been signed by the research participants or their relatives.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
MILLER K D, FIDLER-BENAOUDIA M, KEEGAN T H, et al. Cancer statistics for adolescents and young adults, 2020[J]. CA Cancer J Clin, 2020, 70(6): 443-459.
YANG J D, HAINAUT P, GORES G J, et al. A global view of hepatocellular carcinoma: trends, risk, prevention and management[J]. Nat Rev Gastroenterol Hepatol, 2019, 16(10): 589-604.
ZHU J Y, TANG B F, LI J, et al. Identification and validation of the angiogenic genes for constructing diagnostic, prognostic, and recurrence models for hepatocellular carcinoma[J]. Aging, 2020, 12(9): 7848-7873.
DING Z B, ERICKSEN R E, LEE Q Y, et al. Reprogramming of mitochondrial proline metabolism promotes liver tumorigenesis[J]. Amino Acids, 2021, 53(12): 1807-1815.
SONG B S, MOON J S, TIAN J W, et al. Mitoribosomal defects aggravate liver cancer via aberrant glycolytic flux and T cell exhaustion[J]. J Immunother Cancer, 2022, 10(5): e004337.
LI X J, LI Y M, XU A J, et al. Apoptosis-induced translocation of centromere protein F in its corresponding autoantibody production in hepatocellular carcinoma[J]. Oncoimmunology, 2021, 10(1): 1992104.
CAI Y L, LIN Y X, XIONG X Z, et al. Knockdown expression of MECR, a novel gene of mitochondrial FAS Ⅱ inhibits growth and colony-formation, promotes apoptosis of hepatocelluar carcinoma cells[J]. Biosci Trends, 2019, 13(3): 234-244.
VAZQUEZ F, LIM J H, CHIM H, et al. PGC1α expression defines a subset of human melanoma tumors with increased mitochondrial capacity and resistance to oxidative stress[J]. Cancer Cell, 2013, 23(3): 287-301.
FARGE T, SALAND E, DE TONI F, et al. Chemotherapy-resistant human acute myeloid leukemia cells are not enriched for leukemic stem cells but require oxidative metabolism[J]. Cancer Discov, 2017, 7(7): 716-735.
ABBAS S, LUGTHART S, KAVELAARS F G, et al. Acquired mutations in the genes encoding IDH1 and IDH2 both are recurrent aberrations in acute myeloid leukemia: prevalence and prognostic value[J]. Blood, 2010, 116(12): 2122-2126.
PASINI B, MCWHINNEY S R, BEI T, et al. Clinical and molecular genetics of patients with the Carney-Stratakis syndrome and germline mutations of the genes coding for the succinate dehydrogenase subunits SDHB, SDHC, and SDHD[J]. Eur J Hum Genet, 2008, 16(1): 79-88.
LEHTONEN H J, BLANCO I, PIULATS J M, et al. Conventional renal cancer in a patient with fumarate hydratase mutation[J]. Hum Pathol, 2007, 38(5): 793-796.
FU J, LIU G X, ZHANG X, et al. TRPM8 promotes hepatocellular carcinoma progression by inducing SNORA55 mediated nuclear-mitochondrial communication[J]. Cancer Gene Ther, 2023, 30(5): 738-751.
CHELLA KRISHNAN K, KURT Z, BARRERE-CAIN R, et al. Integration of multi-omics data from mouse diversity panel highlights mitochondrial dysfunction in non-alcoholic fatty liver disease[J]. Cell Syst, 2018, 6(1): 103-115.e7.
GAO F, LIU Q C, LI G P, et al. Identification of ubiquinol cytochrome c reductase hinge (UQCRH) as a potential diagnostic biomarker for lung adenocarcinoma[J]. Open Biol, 2016, 6(6): 150256.
PARK E R, KIM S B, LEE J S, et al. The mitochondrial hinge protein, UQCRH, is a novel prognostic factor for hepatocellular carcinoma[J]. Cancer Med, 2017, 6(4): 749-760.
SUN H, WANG F C, HUANG Y Q, et al. Targeted inhibition of ACLY expression to reverse the resistance of sorafenib in hepatocellular carcinoma[J]. J Cancer, 2022, 13(3): 951-964.
GRANCHI C. ATP citrate lyase (ACLY) inhibitors: an anti-cancer strategy at the crossroads of glucose and lipid metabolism[J]. Eur J Med Chem, 2018, 157: 1276-1291.
HAN Q, CHEN C A, YANG W, et al. ATP-citrate lyase regulates stemness and metastasis in hepatocellular carcinoma via the Wnt/β-catenin signaling pathway[J]. Hepatobiliary Pancreat Dis Int, 2021, 20(3): 251-261.
LEITHNER K, HRZENJAK A, TRÖTZMÜLLER M, et al. PCK2 activation mediates an adaptive response to glucose depletion in lung cancer[J]. Oncogene, 2015, 34(8): 1044-1050.
HYROŠŠOVÁ P, ARAGÓ M, MORENO-FELICI J, et al. PEPCK-M recoups tumor cell anabolic potential in a PKC-ζ-dependent manner[J]. Cancer Metab, 2021, 9(1): 1.
BLUEMEL G, PLANQUE M, MADREITER-SOKOLOWSKI C T, et al. PCK2 opposes mitochondrial respiration and maintains the redox balance in starved lung cancer cells[J]. Free Radic Biol Med, 2021, 176: 34-45.
LIU M X, JIN L, SUN S J, et al. Metabolic reprogramming by PCK1 promotes TCA cataplerosis, oxidative stress and apoptosis in liver cancer cells and suppresses hepatocellular carcinoma[J]. Oncogene, 2018, 37(12): 1637-1653.
COSENTINO K, HERTLEIN V, JENNER A, et al. The interplay between BAX and BAK tunes apoptotic pore growth to control mitochondrial-DNA-mediated inflammation[J]. Mol Cell, 2022, 82(5): 933-949.e9.
LUO X, O'NEILL K L, HUANG K. The third model of Bax/Bak activation: a Bcl-2 family feud finally resolved?[J]. F1000Research, 2020, 9: 935.
HAO B B, LI X J, JIA X L, et al. The novel cereblon modulator CC-885 inhibits mitophagy via selective degradation of BNIP3L[J]. Acta Pharmacol Sin, 2020, 41(9): 1246-1254.
LI Y, ZHENG W Q, LU Y Y, et al. BNIP3L/NIX-mediated mitophagy: molecular mechanisms and implications for human disease[J]. Cell Death Dis, 2021, 13(1): 14.
CHEN Y Y, WANG W H, CHE L, et al. BNIP3L-dependent mitophagy promotes HBx-induced cancer stemness of hepatocellular carcinoma cells via glycolysis metabolism reprogramming[J]. Cancers, 2020, 12(3): 655.
... 在全球范围内,原发性肝癌是最具侵袭性和难以治疗的恶性肿瘤之一[1].肝细胞癌(hepatocellular carcinoma,HCC)约占所有原发性肝癌病例的90%[2].尽管早期和晚期HCC的治疗方法都有所改善[3-4],但近几十年来,许多国家的发病率和死亡率仍然很高[5].由于预后不良,HCC目前是全球第四大常见的癌症相关死亡原因,5年生存率低于21%[6].因此,建立一个能够准确预测HCC预后的模型对于改善HCC预后至关重要.虽然许多分类系统,如美国癌症联合委员会(American Joint Committee on Cancer,AJCC)的TNM分期(American Joint Committee on Cancer-Tumor Node Metastasis Staging)系统、巴塞罗那临床肝癌(Barcelona Clinic Liver Cancer,BCLC)分期和意大利肝癌评分(Cancer of the Liver Italian Program,CLIP)系统,已广泛应用于临床评价HCC患者的预后,但这些分类系统主要是基于临床病理特征,没有考虑分子标志物的重要预后作用[7]. ...
1
... 在全球范围内,原发性肝癌是最具侵袭性和难以治疗的恶性肿瘤之一[1].肝细胞癌(hepatocellular carcinoma,HCC)约占所有原发性肝癌病例的90%[2].尽管早期和晚期HCC的治疗方法都有所改善[3-4],但近几十年来,许多国家的发病率和死亡率仍然很高[5].由于预后不良,HCC目前是全球第四大常见的癌症相关死亡原因,5年生存率低于21%[6].因此,建立一个能够准确预测HCC预后的模型对于改善HCC预后至关重要.虽然许多分类系统,如美国癌症联合委员会(American Joint Committee on Cancer,AJCC)的TNM分期(American Joint Committee on Cancer-Tumor Node Metastasis Staging)系统、巴塞罗那临床肝癌(Barcelona Clinic Liver Cancer,BCLC)分期和意大利肝癌评分(Cancer of the Liver Italian Program,CLIP)系统,已广泛应用于临床评价HCC患者的预后,但这些分类系统主要是基于临床病理特征,没有考虑分子标志物的重要预后作用[7]. ...
1
... 在全球范围内,原发性肝癌是最具侵袭性和难以治疗的恶性肿瘤之一[1].肝细胞癌(hepatocellular carcinoma,HCC)约占所有原发性肝癌病例的90%[2].尽管早期和晚期HCC的治疗方法都有所改善[3-4],但近几十年来,许多国家的发病率和死亡率仍然很高[5].由于预后不良,HCC目前是全球第四大常见的癌症相关死亡原因,5年生存率低于21%[6].因此,建立一个能够准确预测HCC预后的模型对于改善HCC预后至关重要.虽然许多分类系统,如美国癌症联合委员会(American Joint Committee on Cancer,AJCC)的TNM分期(American Joint Committee on Cancer-Tumor Node Metastasis Staging)系统、巴塞罗那临床肝癌(Barcelona Clinic Liver Cancer,BCLC)分期和意大利肝癌评分(Cancer of the Liver Italian Program,CLIP)系统,已广泛应用于临床评价HCC患者的预后,但这些分类系统主要是基于临床病理特征,没有考虑分子标志物的重要预后作用[7]. ...
1
... 在全球范围内,原发性肝癌是最具侵袭性和难以治疗的恶性肿瘤之一[1].肝细胞癌(hepatocellular carcinoma,HCC)约占所有原发性肝癌病例的90%[2].尽管早期和晚期HCC的治疗方法都有所改善[3-4],但近几十年来,许多国家的发病率和死亡率仍然很高[5].由于预后不良,HCC目前是全球第四大常见的癌症相关死亡原因,5年生存率低于21%[6].因此,建立一个能够准确预测HCC预后的模型对于改善HCC预后至关重要.虽然许多分类系统,如美国癌症联合委员会(American Joint Committee on Cancer,AJCC)的TNM分期(American Joint Committee on Cancer-Tumor Node Metastasis Staging)系统、巴塞罗那临床肝癌(Barcelona Clinic Liver Cancer,BCLC)分期和意大利肝癌评分(Cancer of the Liver Italian Program,CLIP)系统,已广泛应用于临床评价HCC患者的预后,但这些分类系统主要是基于临床病理特征,没有考虑分子标志物的重要预后作用[7]. ...
1
... 在全球范围内,原发性肝癌是最具侵袭性和难以治疗的恶性肿瘤之一[1].肝细胞癌(hepatocellular carcinoma,HCC)约占所有原发性肝癌病例的90%[2].尽管早期和晚期HCC的治疗方法都有所改善[3-4],但近几十年来,许多国家的发病率和死亡率仍然很高[5].由于预后不良,HCC目前是全球第四大常见的癌症相关死亡原因,5年生存率低于21%[6].因此,建立一个能够准确预测HCC预后的模型对于改善HCC预后至关重要.虽然许多分类系统,如美国癌症联合委员会(American Joint Committee on Cancer,AJCC)的TNM分期(American Joint Committee on Cancer-Tumor Node Metastasis Staging)系统、巴塞罗那临床肝癌(Barcelona Clinic Liver Cancer,BCLC)分期和意大利肝癌评分(Cancer of the Liver Italian Program,CLIP)系统,已广泛应用于临床评价HCC患者的预后,但这些分类系统主要是基于临床病理特征,没有考虑分子标志物的重要预后作用[7]. ...
1
... 在全球范围内,原发性肝癌是最具侵袭性和难以治疗的恶性肿瘤之一[1].肝细胞癌(hepatocellular carcinoma,HCC)约占所有原发性肝癌病例的90%[2].尽管早期和晚期HCC的治疗方法都有所改善[3-4],但近几十年来,许多国家的发病率和死亡率仍然很高[5].由于预后不良,HCC目前是全球第四大常见的癌症相关死亡原因,5年生存率低于21%[6].因此,建立一个能够准确预测HCC预后的模型对于改善HCC预后至关重要.虽然许多分类系统,如美国癌症联合委员会(American Joint Committee on Cancer,AJCC)的TNM分期(American Joint Committee on Cancer-Tumor Node Metastasis Staging)系统、巴塞罗那临床肝癌(Barcelona Clinic Liver Cancer,BCLC)分期和意大利肝癌评分(Cancer of the Liver Italian Program,CLIP)系统,已广泛应用于临床评价HCC患者的预后,但这些分类系统主要是基于临床病理特征,没有考虑分子标志物的重要预后作用[7]. ...
1
... 在全球范围内,原发性肝癌是最具侵袭性和难以治疗的恶性肿瘤之一[1].肝细胞癌(hepatocellular carcinoma,HCC)约占所有原发性肝癌病例的90%[2].尽管早期和晚期HCC的治疗方法都有所改善[3-4],但近几十年来,许多国家的发病率和死亡率仍然很高[5].由于预后不良,HCC目前是全球第四大常见的癌症相关死亡原因,5年生存率低于21%[6].因此,建立一个能够准确预测HCC预后的模型对于改善HCC预后至关重要.虽然许多分类系统,如美国癌症联合委员会(American Joint Committee on Cancer,AJCC)的TNM分期(American Joint Committee on Cancer-Tumor Node Metastasis Staging)系统、巴塞罗那临床肝癌(Barcelona Clinic Liver Cancer,BCLC)分期和意大利肝癌评分(Cancer of the Liver Italian Program,CLIP)系统,已广泛应用于临床评价HCC患者的预后,但这些分类系统主要是基于临床病理特征,没有考虑分子标志物的重要预后作用[7]. ...
... 本研究着重研究OXPHOS、TCA循环和细胞凋亡这3个通路中的预后相关基因,对95个与HCC预后相关的deNEMG和参与3种有丝分裂通路的NEMG取交集,发现仅6个参与3种有丝分裂通路的deNEMG(UQCRH、ACLY、PCK2、BAK1、BAX和BNIP3L)与HCC的预后密切相关.因此本研究利用这6个基因来构建基于NEMG的风险模型.在这6个NEMG中,UQCRH编码一个定位于线粒体和细胞核的铰链蛋白.UQCRH作为泛素-细胞色素C还原酶复合物(也称为OXPHOS通路的复合物Ⅲ)的主要亚基之一,负责复合物Ⅲ的细胞色素C和C1之间的电子转移.事实上,在肺腺癌[27]和HCC[28]中已经有关于UQCRH表达上调的报道.此外,UQCRH的表达与肿瘤大小、分化程度和有无血管侵袭相关,并与HCC患者的不良预后密切相关[28].ACLY和PCK2参与TCA循环,其中ACLY形成一种细胞质同源四聚体酶,催化TCA循环中的中间体柠檬酸盐转化为草酰乙酸和乙酰辅酶A[29].由于乙酰辅酶A是新脂肪生成和胆固醇生物合成的重要底物之一,因此ACLY是一种重要的脂肪生成酶,在结直肠癌、非小细胞肺癌、乳腺癌、膀胱癌和HCC等癌症类型中高表达[30-32].同时,在HCC患者中,ACLY的高表达与不良预后、肿瘤干性和转移相关[33].PCK2编码一种线粒体磷酸烯醇式丙酮酸羧激酶,该酶催化TCA循环衍生的草酰乙酸转化为磷酸烯醇式丙酮酸,并伴随GTP水解.PCK2在某些癌症类型,如肺癌、前列腺癌、乳腺癌、宫颈癌和睾丸癌中经常上调和激活[34-35],可能以此对抗肿瘤微环境中的代谢饥饿状态[36].此外,既往研究[37]表明,PCK2的下调与HCC的预后和患癌时间有关,并且PCK2的异位过表达可能通过诱导HCC细胞的葡萄糖缺乏而降低细胞活力.BAX、BAK1和BNIP3L是凋亡相关基因,其中BAX和BAK1是线粒体外膜通透性的决定性关键因素[38-40].BNIP3L是线粒体吞噬的双重调节因子,而线粒体吞噬是清除受损线粒体的过程[41].据报道BNIP3L与BAX和BAK相互作用,导致线粒体外膜通透性增强和凋亡加快[42].同时,BNIP3L介导的线粒体吞噬通过糖酵解代谢重编程,促进乙型肝炎病毒X蛋白(hepatitis B virus X protein,HBx)诱导的HCC的肿瘤干性[42]. ...
2
... 本研究着重研究OXPHOS、TCA循环和细胞凋亡这3个通路中的预后相关基因,对95个与HCC预后相关的deNEMG和参与3种有丝分裂通路的NEMG取交集,发现仅6个参与3种有丝分裂通路的deNEMG(UQCRH、ACLY、PCK2、BAK1、BAX和BNIP3L)与HCC的预后密切相关.因此本研究利用这6个基因来构建基于NEMG的风险模型.在这6个NEMG中,UQCRH编码一个定位于线粒体和细胞核的铰链蛋白.UQCRH作为泛素-细胞色素C还原酶复合物(也称为OXPHOS通路的复合物Ⅲ)的主要亚基之一,负责复合物Ⅲ的细胞色素C和C1之间的电子转移.事实上,在肺腺癌[27]和HCC[28]中已经有关于UQCRH表达上调的报道.此外,UQCRH的表达与肿瘤大小、分化程度和有无血管侵袭相关,并与HCC患者的不良预后密切相关[28].ACLY和PCK2参与TCA循环,其中ACLY形成一种细胞质同源四聚体酶,催化TCA循环中的中间体柠檬酸盐转化为草酰乙酸和乙酰辅酶A[29].由于乙酰辅酶A是新脂肪生成和胆固醇生物合成的重要底物之一,因此ACLY是一种重要的脂肪生成酶,在结直肠癌、非小细胞肺癌、乳腺癌、膀胱癌和HCC等癌症类型中高表达[30-32].同时,在HCC患者中,ACLY的高表达与不良预后、肿瘤干性和转移相关[33].PCK2编码一种线粒体磷酸烯醇式丙酮酸羧激酶,该酶催化TCA循环衍生的草酰乙酸转化为磷酸烯醇式丙酮酸,并伴随GTP水解.PCK2在某些癌症类型,如肺癌、前列腺癌、乳腺癌、宫颈癌和睾丸癌中经常上调和激活[34-35],可能以此对抗肿瘤微环境中的代谢饥饿状态[36].此外,既往研究[37]表明,PCK2的下调与HCC的预后和患癌时间有关,并且PCK2的异位过表达可能通过诱导HCC细胞的葡萄糖缺乏而降低细胞活力.BAX、BAK1和BNIP3L是凋亡相关基因,其中BAX和BAK1是线粒体外膜通透性的决定性关键因素[38-40].BNIP3L是线粒体吞噬的双重调节因子,而线粒体吞噬是清除受损线粒体的过程[41].据报道BNIP3L与BAX和BAK相互作用,导致线粒体外膜通透性增强和凋亡加快[42].同时,BNIP3L介导的线粒体吞噬通过糖酵解代谢重编程,促进乙型肝炎病毒X蛋白(hepatitis B virus X protein,HBx)诱导的HCC的肿瘤干性[42]. ...
... [28].ACLY和PCK2参与TCA循环,其中ACLY形成一种细胞质同源四聚体酶,催化TCA循环中的中间体柠檬酸盐转化为草酰乙酸和乙酰辅酶A[29].由于乙酰辅酶A是新脂肪生成和胆固醇生物合成的重要底物之一,因此ACLY是一种重要的脂肪生成酶,在结直肠癌、非小细胞肺癌、乳腺癌、膀胱癌和HCC等癌症类型中高表达[30-32].同时,在HCC患者中,ACLY的高表达与不良预后、肿瘤干性和转移相关[33].PCK2编码一种线粒体磷酸烯醇式丙酮酸羧激酶,该酶催化TCA循环衍生的草酰乙酸转化为磷酸烯醇式丙酮酸,并伴随GTP水解.PCK2在某些癌症类型,如肺癌、前列腺癌、乳腺癌、宫颈癌和睾丸癌中经常上调和激活[34-35],可能以此对抗肿瘤微环境中的代谢饥饿状态[36].此外,既往研究[37]表明,PCK2的下调与HCC的预后和患癌时间有关,并且PCK2的异位过表达可能通过诱导HCC细胞的葡萄糖缺乏而降低细胞活力.BAX、BAK1和BNIP3L是凋亡相关基因,其中BAX和BAK1是线粒体外膜通透性的决定性关键因素[38-40].BNIP3L是线粒体吞噬的双重调节因子,而线粒体吞噬是清除受损线粒体的过程[41].据报道BNIP3L与BAX和BAK相互作用,导致线粒体外膜通透性增强和凋亡加快[42].同时,BNIP3L介导的线粒体吞噬通过糖酵解代谢重编程,促进乙型肝炎病毒X蛋白(hepatitis B virus X protein,HBx)诱导的HCC的肿瘤干性[42]. ...
1
... 本研究着重研究OXPHOS、TCA循环和细胞凋亡这3个通路中的预后相关基因,对95个与HCC预后相关的deNEMG和参与3种有丝分裂通路的NEMG取交集,发现仅6个参与3种有丝分裂通路的deNEMG(UQCRH、ACLY、PCK2、BAK1、BAX和BNIP3L)与HCC的预后密切相关.因此本研究利用这6个基因来构建基于NEMG的风险模型.在这6个NEMG中,UQCRH编码一个定位于线粒体和细胞核的铰链蛋白.UQCRH作为泛素-细胞色素C还原酶复合物(也称为OXPHOS通路的复合物Ⅲ)的主要亚基之一,负责复合物Ⅲ的细胞色素C和C1之间的电子转移.事实上,在肺腺癌[27]和HCC[28]中已经有关于UQCRH表达上调的报道.此外,UQCRH的表达与肿瘤大小、分化程度和有无血管侵袭相关,并与HCC患者的不良预后密切相关[28].ACLY和PCK2参与TCA循环,其中ACLY形成一种细胞质同源四聚体酶,催化TCA循环中的中间体柠檬酸盐转化为草酰乙酸和乙酰辅酶A[29].由于乙酰辅酶A是新脂肪生成和胆固醇生物合成的重要底物之一,因此ACLY是一种重要的脂肪生成酶,在结直肠癌、非小细胞肺癌、乳腺癌、膀胱癌和HCC等癌症类型中高表达[30-32].同时,在HCC患者中,ACLY的高表达与不良预后、肿瘤干性和转移相关[33].PCK2编码一种线粒体磷酸烯醇式丙酮酸羧激酶,该酶催化TCA循环衍生的草酰乙酸转化为磷酸烯醇式丙酮酸,并伴随GTP水解.PCK2在某些癌症类型,如肺癌、前列腺癌、乳腺癌、宫颈癌和睾丸癌中经常上调和激活[34-35],可能以此对抗肿瘤微环境中的代谢饥饿状态[36].此外,既往研究[37]表明,PCK2的下调与HCC的预后和患癌时间有关,并且PCK2的异位过表达可能通过诱导HCC细胞的葡萄糖缺乏而降低细胞活力.BAX、BAK1和BNIP3L是凋亡相关基因,其中BAX和BAK1是线粒体外膜通透性的决定性关键因素[38-40].BNIP3L是线粒体吞噬的双重调节因子,而线粒体吞噬是清除受损线粒体的过程[41].据报道BNIP3L与BAX和BAK相互作用,导致线粒体外膜通透性增强和凋亡加快[42].同时,BNIP3L介导的线粒体吞噬通过糖酵解代谢重编程,促进乙型肝炎病毒X蛋白(hepatitis B virus X protein,HBx)诱导的HCC的肿瘤干性[42]. ...
1
... 本研究着重研究OXPHOS、TCA循环和细胞凋亡这3个通路中的预后相关基因,对95个与HCC预后相关的deNEMG和参与3种有丝分裂通路的NEMG取交集,发现仅6个参与3种有丝分裂通路的deNEMG(UQCRH、ACLY、PCK2、BAK1、BAX和BNIP3L)与HCC的预后密切相关.因此本研究利用这6个基因来构建基于NEMG的风险模型.在这6个NEMG中,UQCRH编码一个定位于线粒体和细胞核的铰链蛋白.UQCRH作为泛素-细胞色素C还原酶复合物(也称为OXPHOS通路的复合物Ⅲ)的主要亚基之一,负责复合物Ⅲ的细胞色素C和C1之间的电子转移.事实上,在肺腺癌[27]和HCC[28]中已经有关于UQCRH表达上调的报道.此外,UQCRH的表达与肿瘤大小、分化程度和有无血管侵袭相关,并与HCC患者的不良预后密切相关[28].ACLY和PCK2参与TCA循环,其中ACLY形成一种细胞质同源四聚体酶,催化TCA循环中的中间体柠檬酸盐转化为草酰乙酸和乙酰辅酶A[29].由于乙酰辅酶A是新脂肪生成和胆固醇生物合成的重要底物之一,因此ACLY是一种重要的脂肪生成酶,在结直肠癌、非小细胞肺癌、乳腺癌、膀胱癌和HCC等癌症类型中高表达[30-32].同时,在HCC患者中,ACLY的高表达与不良预后、肿瘤干性和转移相关[33].PCK2编码一种线粒体磷酸烯醇式丙酮酸羧激酶,该酶催化TCA循环衍生的草酰乙酸转化为磷酸烯醇式丙酮酸,并伴随GTP水解.PCK2在某些癌症类型,如肺癌、前列腺癌、乳腺癌、宫颈癌和睾丸癌中经常上调和激活[34-35],可能以此对抗肿瘤微环境中的代谢饥饿状态[36].此外,既往研究[37]表明,PCK2的下调与HCC的预后和患癌时间有关,并且PCK2的异位过表达可能通过诱导HCC细胞的葡萄糖缺乏而降低细胞活力.BAX、BAK1和BNIP3L是凋亡相关基因,其中BAX和BAK1是线粒体外膜通透性的决定性关键因素[38-40].BNIP3L是线粒体吞噬的双重调节因子,而线粒体吞噬是清除受损线粒体的过程[41].据报道BNIP3L与BAX和BAK相互作用,导致线粒体外膜通透性增强和凋亡加快[42].同时,BNIP3L介导的线粒体吞噬通过糖酵解代谢重编程,促进乙型肝炎病毒X蛋白(hepatitis B virus X protein,HBx)诱导的HCC的肿瘤干性[42]. ...
0
1
... 本研究着重研究OXPHOS、TCA循环和细胞凋亡这3个通路中的预后相关基因,对95个与HCC预后相关的deNEMG和参与3种有丝分裂通路的NEMG取交集,发现仅6个参与3种有丝分裂通路的deNEMG(UQCRH、ACLY、PCK2、BAK1、BAX和BNIP3L)与HCC的预后密切相关.因此本研究利用这6个基因来构建基于NEMG的风险模型.在这6个NEMG中,UQCRH编码一个定位于线粒体和细胞核的铰链蛋白.UQCRH作为泛素-细胞色素C还原酶复合物(也称为OXPHOS通路的复合物Ⅲ)的主要亚基之一,负责复合物Ⅲ的细胞色素C和C1之间的电子转移.事实上,在肺腺癌[27]和HCC[28]中已经有关于UQCRH表达上调的报道.此外,UQCRH的表达与肿瘤大小、分化程度和有无血管侵袭相关,并与HCC患者的不良预后密切相关[28].ACLY和PCK2参与TCA循环,其中ACLY形成一种细胞质同源四聚体酶,催化TCA循环中的中间体柠檬酸盐转化为草酰乙酸和乙酰辅酶A[29].由于乙酰辅酶A是新脂肪生成和胆固醇生物合成的重要底物之一,因此ACLY是一种重要的脂肪生成酶,在结直肠癌、非小细胞肺癌、乳腺癌、膀胱癌和HCC等癌症类型中高表达[30-32].同时,在HCC患者中,ACLY的高表达与不良预后、肿瘤干性和转移相关[33].PCK2编码一种线粒体磷酸烯醇式丙酮酸羧激酶,该酶催化TCA循环衍生的草酰乙酸转化为磷酸烯醇式丙酮酸,并伴随GTP水解.PCK2在某些癌症类型,如肺癌、前列腺癌、乳腺癌、宫颈癌和睾丸癌中经常上调和激活[34-35],可能以此对抗肿瘤微环境中的代谢饥饿状态[36].此外,既往研究[37]表明,PCK2的下调与HCC的预后和患癌时间有关,并且PCK2的异位过表达可能通过诱导HCC细胞的葡萄糖缺乏而降低细胞活力.BAX、BAK1和BNIP3L是凋亡相关基因,其中BAX和BAK1是线粒体外膜通透性的决定性关键因素[38-40].BNIP3L是线粒体吞噬的双重调节因子,而线粒体吞噬是清除受损线粒体的过程[41].据报道BNIP3L与BAX和BAK相互作用,导致线粒体外膜通透性增强和凋亡加快[42].同时,BNIP3L介导的线粒体吞噬通过糖酵解代谢重编程,促进乙型肝炎病毒X蛋白(hepatitis B virus X protein,HBx)诱导的HCC的肿瘤干性[42]. ...
1
... 本研究着重研究OXPHOS、TCA循环和细胞凋亡这3个通路中的预后相关基因,对95个与HCC预后相关的deNEMG和参与3种有丝分裂通路的NEMG取交集,发现仅6个参与3种有丝分裂通路的deNEMG(UQCRH、ACLY、PCK2、BAK1、BAX和BNIP3L)与HCC的预后密切相关.因此本研究利用这6个基因来构建基于NEMG的风险模型.在这6个NEMG中,UQCRH编码一个定位于线粒体和细胞核的铰链蛋白.UQCRH作为泛素-细胞色素C还原酶复合物(也称为OXPHOS通路的复合物Ⅲ)的主要亚基之一,负责复合物Ⅲ的细胞色素C和C1之间的电子转移.事实上,在肺腺癌[27]和HCC[28]中已经有关于UQCRH表达上调的报道.此外,UQCRH的表达与肿瘤大小、分化程度和有无血管侵袭相关,并与HCC患者的不良预后密切相关[28].ACLY和PCK2参与TCA循环,其中ACLY形成一种细胞质同源四聚体酶,催化TCA循环中的中间体柠檬酸盐转化为草酰乙酸和乙酰辅酶A[29].由于乙酰辅酶A是新脂肪生成和胆固醇生物合成的重要底物之一,因此ACLY是一种重要的脂肪生成酶,在结直肠癌、非小细胞肺癌、乳腺癌、膀胱癌和HCC等癌症类型中高表达[30-32].同时,在HCC患者中,ACLY的高表达与不良预后、肿瘤干性和转移相关[33].PCK2编码一种线粒体磷酸烯醇式丙酮酸羧激酶,该酶催化TCA循环衍生的草酰乙酸转化为磷酸烯醇式丙酮酸,并伴随GTP水解.PCK2在某些癌症类型,如肺癌、前列腺癌、乳腺癌、宫颈癌和睾丸癌中经常上调和激活[34-35],可能以此对抗肿瘤微环境中的代谢饥饿状态[36].此外,既往研究[37]表明,PCK2的下调与HCC的预后和患癌时间有关,并且PCK2的异位过表达可能通过诱导HCC细胞的葡萄糖缺乏而降低细胞活力.BAX、BAK1和BNIP3L是凋亡相关基因,其中BAX和BAK1是线粒体外膜通透性的决定性关键因素[38-40].BNIP3L是线粒体吞噬的双重调节因子,而线粒体吞噬是清除受损线粒体的过程[41].据报道BNIP3L与BAX和BAK相互作用,导致线粒体外膜通透性增强和凋亡加快[42].同时,BNIP3L介导的线粒体吞噬通过糖酵解代谢重编程,促进乙型肝炎病毒X蛋白(hepatitis B virus X protein,HBx)诱导的HCC的肿瘤干性[42]. ...
1
... 本研究着重研究OXPHOS、TCA循环和细胞凋亡这3个通路中的预后相关基因,对95个与HCC预后相关的deNEMG和参与3种有丝分裂通路的NEMG取交集,发现仅6个参与3种有丝分裂通路的deNEMG(UQCRH、ACLY、PCK2、BAK1、BAX和BNIP3L)与HCC的预后密切相关.因此本研究利用这6个基因来构建基于NEMG的风险模型.在这6个NEMG中,UQCRH编码一个定位于线粒体和细胞核的铰链蛋白.UQCRH作为泛素-细胞色素C还原酶复合物(也称为OXPHOS通路的复合物Ⅲ)的主要亚基之一,负责复合物Ⅲ的细胞色素C和C1之间的电子转移.事实上,在肺腺癌[27]和HCC[28]中已经有关于UQCRH表达上调的报道.此外,UQCRH的表达与肿瘤大小、分化程度和有无血管侵袭相关,并与HCC患者的不良预后密切相关[28].ACLY和PCK2参与TCA循环,其中ACLY形成一种细胞质同源四聚体酶,催化TCA循环中的中间体柠檬酸盐转化为草酰乙酸和乙酰辅酶A[29].由于乙酰辅酶A是新脂肪生成和胆固醇生物合成的重要底物之一,因此ACLY是一种重要的脂肪生成酶,在结直肠癌、非小细胞肺癌、乳腺癌、膀胱癌和HCC等癌症类型中高表达[30-32].同时,在HCC患者中,ACLY的高表达与不良预后、肿瘤干性和转移相关[33].PCK2编码一种线粒体磷酸烯醇式丙酮酸羧激酶,该酶催化TCA循环衍生的草酰乙酸转化为磷酸烯醇式丙酮酸,并伴随GTP水解.PCK2在某些癌症类型,如肺癌、前列腺癌、乳腺癌、宫颈癌和睾丸癌中经常上调和激活[34-35],可能以此对抗肿瘤微环境中的代谢饥饿状态[36].此外,既往研究[37]表明,PCK2的下调与HCC的预后和患癌时间有关,并且PCK2的异位过表达可能通过诱导HCC细胞的葡萄糖缺乏而降低细胞活力.BAX、BAK1和BNIP3L是凋亡相关基因,其中BAX和BAK1是线粒体外膜通透性的决定性关键因素[38-40].BNIP3L是线粒体吞噬的双重调节因子,而线粒体吞噬是清除受损线粒体的过程[41].据报道BNIP3L与BAX和BAK相互作用,导致线粒体外膜通透性增强和凋亡加快[42].同时,BNIP3L介导的线粒体吞噬通过糖酵解代谢重编程,促进乙型肝炎病毒X蛋白(hepatitis B virus X protein,HBx)诱导的HCC的肿瘤干性[42]. ...
1
... 本研究着重研究OXPHOS、TCA循环和细胞凋亡这3个通路中的预后相关基因,对95个与HCC预后相关的deNEMG和参与3种有丝分裂通路的NEMG取交集,发现仅6个参与3种有丝分裂通路的deNEMG(UQCRH、ACLY、PCK2、BAK1、BAX和BNIP3L)与HCC的预后密切相关.因此本研究利用这6个基因来构建基于NEMG的风险模型.在这6个NEMG中,UQCRH编码一个定位于线粒体和细胞核的铰链蛋白.UQCRH作为泛素-细胞色素C还原酶复合物(也称为OXPHOS通路的复合物Ⅲ)的主要亚基之一,负责复合物Ⅲ的细胞色素C和C1之间的电子转移.事实上,在肺腺癌[27]和HCC[28]中已经有关于UQCRH表达上调的报道.此外,UQCRH的表达与肿瘤大小、分化程度和有无血管侵袭相关,并与HCC患者的不良预后密切相关[28].ACLY和PCK2参与TCA循环,其中ACLY形成一种细胞质同源四聚体酶,催化TCA循环中的中间体柠檬酸盐转化为草酰乙酸和乙酰辅酶A[29].由于乙酰辅酶A是新脂肪生成和胆固醇生物合成的重要底物之一,因此ACLY是一种重要的脂肪生成酶,在结直肠癌、非小细胞肺癌、乳腺癌、膀胱癌和HCC等癌症类型中高表达[30-32].同时,在HCC患者中,ACLY的高表达与不良预后、肿瘤干性和转移相关[33].PCK2编码一种线粒体磷酸烯醇式丙酮酸羧激酶,该酶催化TCA循环衍生的草酰乙酸转化为磷酸烯醇式丙酮酸,并伴随GTP水解.PCK2在某些癌症类型,如肺癌、前列腺癌、乳腺癌、宫颈癌和睾丸癌中经常上调和激活[34-35],可能以此对抗肿瘤微环境中的代谢饥饿状态[36].此外,既往研究[37]表明,PCK2的下调与HCC的预后和患癌时间有关,并且PCK2的异位过表达可能通过诱导HCC细胞的葡萄糖缺乏而降低细胞活力.BAX、BAK1和BNIP3L是凋亡相关基因,其中BAX和BAK1是线粒体外膜通透性的决定性关键因素[38-40].BNIP3L是线粒体吞噬的双重调节因子,而线粒体吞噬是清除受损线粒体的过程[41].据报道BNIP3L与BAX和BAK相互作用,导致线粒体外膜通透性增强和凋亡加快[42].同时,BNIP3L介导的线粒体吞噬通过糖酵解代谢重编程,促进乙型肝炎病毒X蛋白(hepatitis B virus X protein,HBx)诱导的HCC的肿瘤干性[42]. ...
1
... 本研究着重研究OXPHOS、TCA循环和细胞凋亡这3个通路中的预后相关基因,对95个与HCC预后相关的deNEMG和参与3种有丝分裂通路的NEMG取交集,发现仅6个参与3种有丝分裂通路的deNEMG(UQCRH、ACLY、PCK2、BAK1、BAX和BNIP3L)与HCC的预后密切相关.因此本研究利用这6个基因来构建基于NEMG的风险模型.在这6个NEMG中,UQCRH编码一个定位于线粒体和细胞核的铰链蛋白.UQCRH作为泛素-细胞色素C还原酶复合物(也称为OXPHOS通路的复合物Ⅲ)的主要亚基之一,负责复合物Ⅲ的细胞色素C和C1之间的电子转移.事实上,在肺腺癌[27]和HCC[28]中已经有关于UQCRH表达上调的报道.此外,UQCRH的表达与肿瘤大小、分化程度和有无血管侵袭相关,并与HCC患者的不良预后密切相关[28].ACLY和PCK2参与TCA循环,其中ACLY形成一种细胞质同源四聚体酶,催化TCA循环中的中间体柠檬酸盐转化为草酰乙酸和乙酰辅酶A[29].由于乙酰辅酶A是新脂肪生成和胆固醇生物合成的重要底物之一,因此ACLY是一种重要的脂肪生成酶,在结直肠癌、非小细胞肺癌、乳腺癌、膀胱癌和HCC等癌症类型中高表达[30-32].同时,在HCC患者中,ACLY的高表达与不良预后、肿瘤干性和转移相关[33].PCK2编码一种线粒体磷酸烯醇式丙酮酸羧激酶,该酶催化TCA循环衍生的草酰乙酸转化为磷酸烯醇式丙酮酸,并伴随GTP水解.PCK2在某些癌症类型,如肺癌、前列腺癌、乳腺癌、宫颈癌和睾丸癌中经常上调和激活[34-35],可能以此对抗肿瘤微环境中的代谢饥饿状态[36].此外,既往研究[37]表明,PCK2的下调与HCC的预后和患癌时间有关,并且PCK2的异位过表达可能通过诱导HCC细胞的葡萄糖缺乏而降低细胞活力.BAX、BAK1和BNIP3L是凋亡相关基因,其中BAX和BAK1是线粒体外膜通透性的决定性关键因素[38-40].BNIP3L是线粒体吞噬的双重调节因子,而线粒体吞噬是清除受损线粒体的过程[41].据报道BNIP3L与BAX和BAK相互作用,导致线粒体外膜通透性增强和凋亡加快[42].同时,BNIP3L介导的线粒体吞噬通过糖酵解代谢重编程,促进乙型肝炎病毒X蛋白(hepatitis B virus X protein,HBx)诱导的HCC的肿瘤干性[42]. ...
1
... 本研究着重研究OXPHOS、TCA循环和细胞凋亡这3个通路中的预后相关基因,对95个与HCC预后相关的deNEMG和参与3种有丝分裂通路的NEMG取交集,发现仅6个参与3种有丝分裂通路的deNEMG(UQCRH、ACLY、PCK2、BAK1、BAX和BNIP3L)与HCC的预后密切相关.因此本研究利用这6个基因来构建基于NEMG的风险模型.在这6个NEMG中,UQCRH编码一个定位于线粒体和细胞核的铰链蛋白.UQCRH作为泛素-细胞色素C还原酶复合物(也称为OXPHOS通路的复合物Ⅲ)的主要亚基之一,负责复合物Ⅲ的细胞色素C和C1之间的电子转移.事实上,在肺腺癌[27]和HCC[28]中已经有关于UQCRH表达上调的报道.此外,UQCRH的表达与肿瘤大小、分化程度和有无血管侵袭相关,并与HCC患者的不良预后密切相关[28].ACLY和PCK2参与TCA循环,其中ACLY形成一种细胞质同源四聚体酶,催化TCA循环中的中间体柠檬酸盐转化为草酰乙酸和乙酰辅酶A[29].由于乙酰辅酶A是新脂肪生成和胆固醇生物合成的重要底物之一,因此ACLY是一种重要的脂肪生成酶,在结直肠癌、非小细胞肺癌、乳腺癌、膀胱癌和HCC等癌症类型中高表达[30-32].同时,在HCC患者中,ACLY的高表达与不良预后、肿瘤干性和转移相关[33].PCK2编码一种线粒体磷酸烯醇式丙酮酸羧激酶,该酶催化TCA循环衍生的草酰乙酸转化为磷酸烯醇式丙酮酸,并伴随GTP水解.PCK2在某些癌症类型,如肺癌、前列腺癌、乳腺癌、宫颈癌和睾丸癌中经常上调和激活[34-35],可能以此对抗肿瘤微环境中的代谢饥饿状态[36].此外,既往研究[37]表明,PCK2的下调与HCC的预后和患癌时间有关,并且PCK2的异位过表达可能通过诱导HCC细胞的葡萄糖缺乏而降低细胞活力.BAX、BAK1和BNIP3L是凋亡相关基因,其中BAX和BAK1是线粒体外膜通透性的决定性关键因素[38-40].BNIP3L是线粒体吞噬的双重调节因子,而线粒体吞噬是清除受损线粒体的过程[41].据报道BNIP3L与BAX和BAK相互作用,导致线粒体外膜通透性增强和凋亡加快[42].同时,BNIP3L介导的线粒体吞噬通过糖酵解代谢重编程,促进乙型肝炎病毒X蛋白(hepatitis B virus X protein,HBx)诱导的HCC的肿瘤干性[42]. ...
1
... 本研究着重研究OXPHOS、TCA循环和细胞凋亡这3个通路中的预后相关基因,对95个与HCC预后相关的deNEMG和参与3种有丝分裂通路的NEMG取交集,发现仅6个参与3种有丝分裂通路的deNEMG(UQCRH、ACLY、PCK2、BAK1、BAX和BNIP3L)与HCC的预后密切相关.因此本研究利用这6个基因来构建基于NEMG的风险模型.在这6个NEMG中,UQCRH编码一个定位于线粒体和细胞核的铰链蛋白.UQCRH作为泛素-细胞色素C还原酶复合物(也称为OXPHOS通路的复合物Ⅲ)的主要亚基之一,负责复合物Ⅲ的细胞色素C和C1之间的电子转移.事实上,在肺腺癌[27]和HCC[28]中已经有关于UQCRH表达上调的报道.此外,UQCRH的表达与肿瘤大小、分化程度和有无血管侵袭相关,并与HCC患者的不良预后密切相关[28].ACLY和PCK2参与TCA循环,其中ACLY形成一种细胞质同源四聚体酶,催化TCA循环中的中间体柠檬酸盐转化为草酰乙酸和乙酰辅酶A[29].由于乙酰辅酶A是新脂肪生成和胆固醇生物合成的重要底物之一,因此ACLY是一种重要的脂肪生成酶,在结直肠癌、非小细胞肺癌、乳腺癌、膀胱癌和HCC等癌症类型中高表达[30-32].同时,在HCC患者中,ACLY的高表达与不良预后、肿瘤干性和转移相关[33].PCK2编码一种线粒体磷酸烯醇式丙酮酸羧激酶,该酶催化TCA循环衍生的草酰乙酸转化为磷酸烯醇式丙酮酸,并伴随GTP水解.PCK2在某些癌症类型,如肺癌、前列腺癌、乳腺癌、宫颈癌和睾丸癌中经常上调和激活[34-35],可能以此对抗肿瘤微环境中的代谢饥饿状态[36].此外,既往研究[37]表明,PCK2的下调与HCC的预后和患癌时间有关,并且PCK2的异位过表达可能通过诱导HCC细胞的葡萄糖缺乏而降低细胞活力.BAX、BAK1和BNIP3L是凋亡相关基因,其中BAX和BAK1是线粒体外膜通透性的决定性关键因素[38-40].BNIP3L是线粒体吞噬的双重调节因子,而线粒体吞噬是清除受损线粒体的过程[41].据报道BNIP3L与BAX和BAK相互作用,导致线粒体外膜通透性增强和凋亡加快[42].同时,BNIP3L介导的线粒体吞噬通过糖酵解代谢重编程,促进乙型肝炎病毒X蛋白(hepatitis B virus X protein,HBx)诱导的HCC的肿瘤干性[42]. ...
0
1
... 本研究着重研究OXPHOS、TCA循环和细胞凋亡这3个通路中的预后相关基因,对95个与HCC预后相关的deNEMG和参与3种有丝分裂通路的NEMG取交集,发现仅6个参与3种有丝分裂通路的deNEMG(UQCRH、ACLY、PCK2、BAK1、BAX和BNIP3L)与HCC的预后密切相关.因此本研究利用这6个基因来构建基于NEMG的风险模型.在这6个NEMG中,UQCRH编码一个定位于线粒体和细胞核的铰链蛋白.UQCRH作为泛素-细胞色素C还原酶复合物(也称为OXPHOS通路的复合物Ⅲ)的主要亚基之一,负责复合物Ⅲ的细胞色素C和C1之间的电子转移.事实上,在肺腺癌[27]和HCC[28]中已经有关于UQCRH表达上调的报道.此外,UQCRH的表达与肿瘤大小、分化程度和有无血管侵袭相关,并与HCC患者的不良预后密切相关[28].ACLY和PCK2参与TCA循环,其中ACLY形成一种细胞质同源四聚体酶,催化TCA循环中的中间体柠檬酸盐转化为草酰乙酸和乙酰辅酶A[29].由于乙酰辅酶A是新脂肪生成和胆固醇生物合成的重要底物之一,因此ACLY是一种重要的脂肪生成酶,在结直肠癌、非小细胞肺癌、乳腺癌、膀胱癌和HCC等癌症类型中高表达[30-32].同时,在HCC患者中,ACLY的高表达与不良预后、肿瘤干性和转移相关[33].PCK2编码一种线粒体磷酸烯醇式丙酮酸羧激酶,该酶催化TCA循环衍生的草酰乙酸转化为磷酸烯醇式丙酮酸,并伴随GTP水解.PCK2在某些癌症类型,如肺癌、前列腺癌、乳腺癌、宫颈癌和睾丸癌中经常上调和激活[34-35],可能以此对抗肿瘤微环境中的代谢饥饿状态[36].此外,既往研究[37]表明,PCK2的下调与HCC的预后和患癌时间有关,并且PCK2的异位过表达可能通过诱导HCC细胞的葡萄糖缺乏而降低细胞活力.BAX、BAK1和BNIP3L是凋亡相关基因,其中BAX和BAK1是线粒体外膜通透性的决定性关键因素[38-40].BNIP3L是线粒体吞噬的双重调节因子,而线粒体吞噬是清除受损线粒体的过程[41].据报道BNIP3L与BAX和BAK相互作用,导致线粒体外膜通透性增强和凋亡加快[42].同时,BNIP3L介导的线粒体吞噬通过糖酵解代谢重编程,促进乙型肝炎病毒X蛋白(hepatitis B virus X protein,HBx)诱导的HCC的肿瘤干性[42]. ...
1
... 本研究着重研究OXPHOS、TCA循环和细胞凋亡这3个通路中的预后相关基因,对95个与HCC预后相关的deNEMG和参与3种有丝分裂通路的NEMG取交集,发现仅6个参与3种有丝分裂通路的deNEMG(UQCRH、ACLY、PCK2、BAK1、BAX和BNIP3L)与HCC的预后密切相关.因此本研究利用这6个基因来构建基于NEMG的风险模型.在这6个NEMG中,UQCRH编码一个定位于线粒体和细胞核的铰链蛋白.UQCRH作为泛素-细胞色素C还原酶复合物(也称为OXPHOS通路的复合物Ⅲ)的主要亚基之一,负责复合物Ⅲ的细胞色素C和C1之间的电子转移.事实上,在肺腺癌[27]和HCC[28]中已经有关于UQCRH表达上调的报道.此外,UQCRH的表达与肿瘤大小、分化程度和有无血管侵袭相关,并与HCC患者的不良预后密切相关[28].ACLY和PCK2参与TCA循环,其中ACLY形成一种细胞质同源四聚体酶,催化TCA循环中的中间体柠檬酸盐转化为草酰乙酸和乙酰辅酶A[29].由于乙酰辅酶A是新脂肪生成和胆固醇生物合成的重要底物之一,因此ACLY是一种重要的脂肪生成酶,在结直肠癌、非小细胞肺癌、乳腺癌、膀胱癌和HCC等癌症类型中高表达[30-32].同时,在HCC患者中,ACLY的高表达与不良预后、肿瘤干性和转移相关[33].PCK2编码一种线粒体磷酸烯醇式丙酮酸羧激酶,该酶催化TCA循环衍生的草酰乙酸转化为磷酸烯醇式丙酮酸,并伴随GTP水解.PCK2在某些癌症类型,如肺癌、前列腺癌、乳腺癌、宫颈癌和睾丸癌中经常上调和激活[34-35],可能以此对抗肿瘤微环境中的代谢饥饿状态[36].此外,既往研究[37]表明,PCK2的下调与HCC的预后和患癌时间有关,并且PCK2的异位过表达可能通过诱导HCC细胞的葡萄糖缺乏而降低细胞活力.BAX、BAK1和BNIP3L是凋亡相关基因,其中BAX和BAK1是线粒体外膜通透性的决定性关键因素[38-40].BNIP3L是线粒体吞噬的双重调节因子,而线粒体吞噬是清除受损线粒体的过程[41].据报道BNIP3L与BAX和BAK相互作用,导致线粒体外膜通透性增强和凋亡加快[42].同时,BNIP3L介导的线粒体吞噬通过糖酵解代谢重编程,促进乙型肝炎病毒X蛋白(hepatitis B virus X protein,HBx)诱导的HCC的肿瘤干性[42]. ...
2
... 本研究着重研究OXPHOS、TCA循环和细胞凋亡这3个通路中的预后相关基因,对95个与HCC预后相关的deNEMG和参与3种有丝分裂通路的NEMG取交集,发现仅6个参与3种有丝分裂通路的deNEMG(UQCRH、ACLY、PCK2、BAK1、BAX和BNIP3L)与HCC的预后密切相关.因此本研究利用这6个基因来构建基于NEMG的风险模型.在这6个NEMG中,UQCRH编码一个定位于线粒体和细胞核的铰链蛋白.UQCRH作为泛素-细胞色素C还原酶复合物(也称为OXPHOS通路的复合物Ⅲ)的主要亚基之一,负责复合物Ⅲ的细胞色素C和C1之间的电子转移.事实上,在肺腺癌[27]和HCC[28]中已经有关于UQCRH表达上调的报道.此外,UQCRH的表达与肿瘤大小、分化程度和有无血管侵袭相关,并与HCC患者的不良预后密切相关[28].ACLY和PCK2参与TCA循环,其中ACLY形成一种细胞质同源四聚体酶,催化TCA循环中的中间体柠檬酸盐转化为草酰乙酸和乙酰辅酶A[29].由于乙酰辅酶A是新脂肪生成和胆固醇生物合成的重要底物之一,因此ACLY是一种重要的脂肪生成酶,在结直肠癌、非小细胞肺癌、乳腺癌、膀胱癌和HCC等癌症类型中高表达[30-32].同时,在HCC患者中,ACLY的高表达与不良预后、肿瘤干性和转移相关[33].PCK2编码一种线粒体磷酸烯醇式丙酮酸羧激酶,该酶催化TCA循环衍生的草酰乙酸转化为磷酸烯醇式丙酮酸,并伴随GTP水解.PCK2在某些癌症类型,如肺癌、前列腺癌、乳腺癌、宫颈癌和睾丸癌中经常上调和激活[34-35],可能以此对抗肿瘤微环境中的代谢饥饿状态[36].此外,既往研究[37]表明,PCK2的下调与HCC的预后和患癌时间有关,并且PCK2的异位过表达可能通过诱导HCC细胞的葡萄糖缺乏而降低细胞活力.BAX、BAK1和BNIP3L是凋亡相关基因,其中BAX和BAK1是线粒体外膜通透性的决定性关键因素[38-40].BNIP3L是线粒体吞噬的双重调节因子,而线粒体吞噬是清除受损线粒体的过程[41].据报道BNIP3L与BAX和BAK相互作用,导致线粒体外膜通透性增强和凋亡加快[42].同时,BNIP3L介导的线粒体吞噬通过糖酵解代谢重编程,促进乙型肝炎病毒X蛋白(hepatitis B virus X protein,HBx)诱导的HCC的肿瘤干性[42]. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}