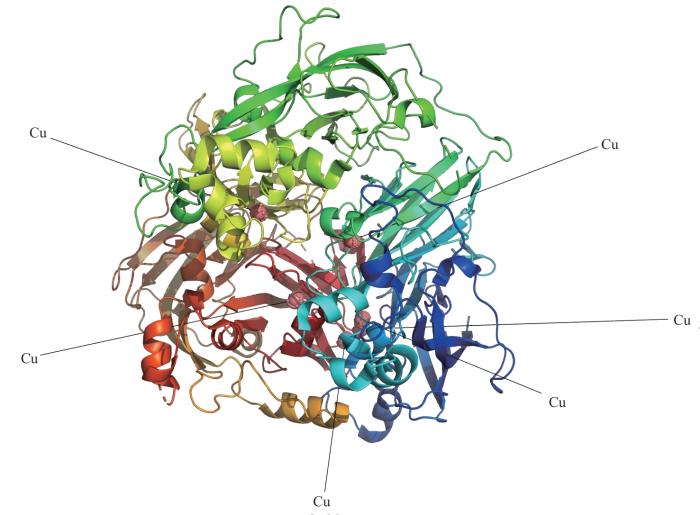
Ceruloplasmin (Cp) is a crucial protein secreted by the liver and plays a vital role in regulating the distribution and transport of copper throughout the body, thereby maintaining copper homeostasis. Additionally, Cp functions as a significant enzyme known as ferroxidase, which is involved in iron metabolism within the body. Numerous studies have suggested a close relationship between Cp and metabolic disorders, such as diabetes and cardiovascular diseases. Recent research has also shed light on the involvement of Cp in the regulation of lipid metabolism. The various activities associated with lipid metabolism, including lipid synthesis, adipose hydrolysis, fatty acid oxidation, lipid transport, and absorption, collectively contribute to maintaining lipid homeostasis. Dysregulation of lipid metabolism can lead to metabolic disorders and cardiovascular complications. Cp regulates lipid metabolism through two main mechanisms. Firstly, Cp participates in the regulation of oxidative stress by modulating iron metabolism through its ferroxidase activity and involvement in redox reaction. Secondly, copper along with copper-dependent enzymes directly participates in the processes such as cholesterol metabolism, lipoprotein metabolism, and fatty acid synthesis. As a result, the role of Cp in maintaining the homeostasis of copper and iron allows it to regulate lipid metabolism by influencing copper or iron-dependent enzymes and related pathways. Although the correlation between Cp and lipid metabolism has been identified, an in-depth exploration of the precise mechanisms by which Cp governs lipid metabolism is warranted. This article provides an overview of the role of Cp in lipid metabolism and highlights the progress in related research, with the aim of providing new insights for the development and treatment of disorders related to lipid metabolism.
JIANG Quanxin, CHEN Suzhen, LIU Junli. Research progress in ceruloplasmin regulation of lipid metabolism homeostasis. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2024, 44(1): 124-130 doi:10.3969/j.issn.1674-8115.2024.01.014
肝豆状核变性是一种常染色体隐性遗传疾病,其特点是铜在体内的蓄积和中毒。该疾病由ATP7B酶的突变引起,该酶参与排泄过量铜的过程[48]。肝豆状核变性患者肝脏中铜的大量积累表明ATP7B对于从肝脏排泄铜是必不可少的。铜的胆汁排泄是维持生理铜平衡的主要过程,其速率随着肝细胞中铜浓度的升高而增加。作为调节全身铜稳态的中枢器官,肝脏是最早和最常见的因铜毒性积累受到损伤的器官。肝豆状核变性患者的肝脏铜离子浓度通常增加约5倍,病程早期即可检测到肝细胞线粒体完整性和功能损伤,肝脏能量代谢受损和胆固醇生物合成相关基因的下调,上述过程进而导致肝脏脂肪变性、慢性肝细胞损伤[48]。肝豆状核变性患者和动物模型都表现出线粒体脂质过氧化增加。在线粒体磷脂中,心磷脂是一种铜敏感靶标,可因铜产生的自由基积累而改变其组成或酰化状态导致线粒体功能障碍[49]。此外,研究[50]表明,Cp是低密度脂蛋白(low density lipoprotein,LDL)氧化的有效催化剂。在心血管疾病的研究中,高血脂等致病因素可导致血管内皮细胞或血管平滑肌细胞表面的Cp粘连,Cp会释放二价铜离子氧化LDL并促进炎症因子或者活性氧(reactive oxygen species,ROS)的生成,从而导致更严重的血管损伤[51]。
The manuscript was drafted by JIANG Quanxin. CHEN Suzhen and LIU Junli were responsible for the guidance and revision of the paper. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
LU Y, WEI X, CHEN M L, et al. Non-ceruloplasmin-bound copper and copper speciation in serum with extraction using functionalized dendritic silica spheres followed by ICP-MS detection[J]. Anal Chim Acta, 2023, 1251: 340993.
CHEN M, ZHENG J S, LIU G H, et al. Ceruloplasmin and hephaestin jointly protect the exocrine pancreas against oxidative damage by facilitating iron efflux[J]. Redox Biol, 2018, 17: 432-439.
WANG P W, WU T H, PAN T L, et al. Integrated proteome and cytokine profiles reveal ceruloplasmin eliciting liver allograft tolerance via antioxidant cascades[J]. Front Immunol, 2018, 9: 2216.
CHEN J, JIANG Y H, SHI H, et al. The molecular mechanisms of copper metabolism and its roles in human diseases[J]. Pflugers Arch, 2020, 472(10): 1415-1429.
NEŞELIOĞLU S, OĞUZ E F, EREL Ö. Development of a new colorimetric, kinetic and automated ceruloplasmin ferroxidase activity measurement method[J]. Antioxidants (Basel), 2022, 11(11): 2187.
SHANG Y X, LUO M Y, YAO F P, et al. Ceruloplasmin suppresses ferroptosis by regulating iron homeostasis in hepatocellular carcinoma cells[J]. Cell Signal, 2020, 72: 109633.
DOGUER C, HA J H, COLLINS J F. Intersection of iron and copper metabolism in the mammalian intestine and liver[J]. Compr Physiol, 2018, 8(4): 1433-1461.
MARCHI G, BUSTI F, LIRA ZIDANES A, et al. Aceruloplasminemia: a severe neurodegenerative disorder deserving an early diagnosis[J]. Front Neurosci, 2019, 13: 325.
KO C W, QU J, BLACK D D, et al. Regulation of intestinal lipid metabolism: current concepts and relevance to disease[J]. Nat Rev Gastroenterol Hepatol, 2020, 17(3): 169-183.
LEE E, KORF H, VIDAL-PUIG A. An adipocentric perspective on the development and progression of non-alcoholic fatty liver disease[J]. J Hepatol, 2023, 78(5): 1048-1062.
MA Y B, TEMKIN S M, HAWKRIDGE A M, et al. Fatty acid oxidation: an emerging facet of metabolic transformation in cancer[J]. Cancer Lett, 2018, 435: 92-100.
ZHONG S S, LI L X, SHEN X, et al. An update on lipid oxidation and inflammation in cardiovascular diseases[J]. Free Radic Biol Med, 2019, 144: 266-278.
KIM C H, PARK J Y, KIM J Y, et al. Elevated serum ceruloplasmin levels in subjects with metabolic syndrome: a population-based study[J]. Metabolism, 2002, 51(7): 838-842.
ENGSTRÖM G, STAVENOW L, HEDBLAD B, et al. Inflammation-sensitive plasma proteins, diabetes, and mortality and incidence of myocardial infarction and stroke: a population-based study[J]. Diabetes, 2003, 52(2): 442-447.
XIE L P, YUAN Y M, XU S M, et al. Downregulation of hepatic ceruloplasmin ameliorates NAFLD via SCO1-AMPK-LKB1 complex[J]. Cell Rep, 2022, 41(3): 111498.
GUTHRIE L M, SOMA S, YUAN S, et al. Elesclomol alleviates Menkes pathology and mortality by escorting Cu to cuproenzymes in mice[J]. Science, 2020, 368(6491): 620-625.
AIGNER E, STRASSER M, HAUFE H, et al. A role for low hepatic copper concentrations in nonalcoholic fatty liver disease[J]. Am J Gastroenterol, 2010, 105(9): 1978-1985.
HEFFERN M C, PARK H M, AU-YEUNG H Y, et al. In vivo bioluminescence imaging reveals copper deficiency in a murine model of nonalcoholic fatty liver disease[J]. Proc Natl Acad Sci USA, 2016, 113(50): 14219-14224.
TREFTS E, SHAW R J. AMPK: restoring metabolic homeostasis over space and time[J]. Mol Cell, 2021, 81(18): 3677-3690.
LIN S C, HARDIE D G. AMPK: sensing glucose as well as cellular energy status[J]. Cell Metab, 2018, 27(2): 299-313.
CUSI K, ALKHOURI N, HARRISON S A, et al. Efficacy and safety of PXL770, a direct AMP kinase activator, for the treatment of non-alcoholic fatty liver disease (STAMP-NAFLD): a randomised, double-blind, placebo-controlled, phase 2a study[J]. Lancet Gastroenterol Hepatol, 2021, 6(11): 889-902.
YANG H J, RALLE M, WOLFGANG M J, et al. Copper-dependent amino oxidase 3 governs selection of metabolic fuels in adipocytes[J]. PLoS Biol, 2018, 16(9): e2006519.
RAIA S, CONTI A, ZANARDI A, et al. Ceruloplasmin-deficient mice show dysregulation of lipid metabolism in liver and adipose tissue reduced by a protein replacement[J]. Int J Mol Sci, 2023, 24(2): 1150.
MANNELLA V, CHAABANE L, CANU T, et al. Lipid dysmetabolism in ceruloplasmin-deficient mice revealed both in vivo and ex vivo by MRI, MRS and NMR analyses[J]. FEBS Open Bio, 2023. DOI: 10.1002/2211-5463.13740.
LIU Z D, WANG M, ZHANG C B, et al. Molecular functions of ceruloplasmin in metabolic disease pathology[J]. Diabetes Metab Syndr Obes, 2022, 15: 695-711.
CORRADINI E, BUZZETTI E, DONGIOVANNI P, et al. Ceruloplasmin gene variants are associated with hyperferritinemia and increased liver iron in patients with NAFLD[J]. J Hepatol, 2021, 75(3): 506-513.
THEPSUWAN P, BHATTACHARYA A, SONG Z F, et al. Hepatic SEL1L-HRD1 ER-associated degradation regulates systemic iron homeostasis via ceruloplasmin[J]. Proc Natl Acad Sci USA, 2023, 120(2): e2212644120.
PATEL B N, DUNN R J, JEONG S Y, et al. Ceruloplasmin regulates iron levels in the CNS and prevents free radical injury[J]. J Neurosci, 2002, 22(15): 6578-6586.
YURKOVA I L, ARNHOLD J, FITZL G, et al. Fragmentation of mitochondrial cardiolipin by copper ions in the Atp7b-/- mouse model of Wilson′s disease[J]. Chem Phys Lipids, 2011, 164(5): 393-400.
ARENAS DE LARRIVA A P, LIMIA-PÉREZ L, ALCALÁ-DÍAZ J F, et al. Ceruloplasmin and coronary heart disease: a systematic review[J]. Nutrients, 2020, 12(10): 3219.
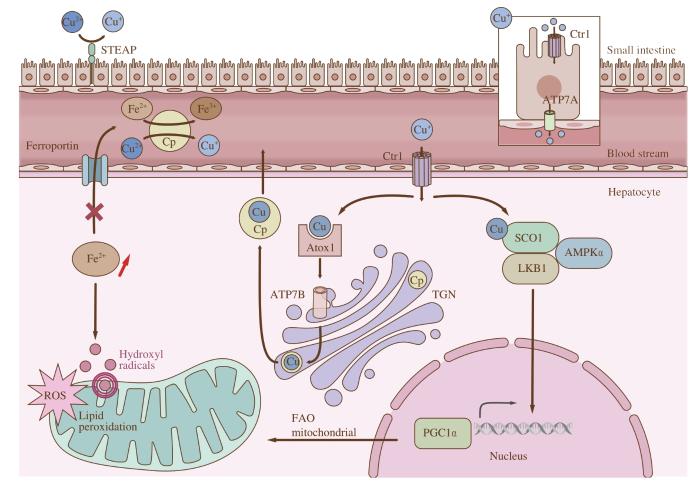
... 由于Cp能携带肝脏的铜分布到机体的不同组织中,因此其在血中的水平升高可相应地引起肝脏中铜分布的不足.我们课题组之前的研究[29]发现,NAFLD患者血清中Cp水平上调导致肝细胞内的铜离子输出增多,从而引起肝脏铜含量降低.肝脏中铜含量的变化会引发一些疾病,如门克斯病(Menkes disease);该疾病是由ATP7A基因突变引起,会导致不同组织中铜的缺乏[30].此外,AIGNER等[31]的研究表明,NAFLD患者的肝脏铜浓度明显低于健康人群.HEFFERN等[32]使用Copper-Caged Luciferin-1(一种生物发光报告基因)进行体内组织特异性铜成像,监测活体动物体内的铜水平;他们也发现,在饮食诱导的NAFLD小鼠模型中,肝脏铜离子水平显著降低.TANG等[33]的研究表明,膳食铜离子缺乏会导致肝脂肪酸合成酶基因转录速度加快,从而提高肝脂质合成速度.核转录因子甾醇调节元件结合蛋白-1(sterol regulatory element-binding protein 1,SREBP-1)是脂肪酸合成的重要转录因子.在铜离子不足的情况下,成熟的SREBP-1会迁移到细胞核,从而促进脂肪酸从头合成的关键酶基因的转录. ...
... 我们课题组之前的研究[29]表明,Cp在调节肝脏TAG代谢中起到重要作用,并通过影响AMP依赖的蛋白激酶(adenosine 5′-monophosphate activated protein kinase,AMPK)信号通路发挥作用.AMPK是一种能量开关蛋白,调控细胞能量利用过程,包括脂质和葡萄糖代谢等.AMPK可激活过氧化物酶体增殖物活化受体γ共激活因子1α(peroxisome proliferator-activated receptor γ coactivator 1α,PGC1α),并通过与过氧化物酶体增殖物活化受体γ(peroxisome proliferator-activated receptor γ,PPARγ)或雌激素相关受体的相互作用,增加线粒体合成和促进脂肪酸的氧化.因此激活的AMPK可作为NAFLD的治疗靶点之一,通过活化AMPK在肝脏中发挥降脂作用[34-37].临床研究[38]发现,一种AMPK激活剂PXL770在口服给药后12周,可平均减少14.7%的肝脏脂肪含量.此外,我们课题组的研究[29]还发现,抑制肝细胞中Cp的分泌可恢复肝脏铜含量至正常水平,并促进AMPK与细胞色素C氧化酶装配蛋白(synthesis of cytochrome C oxidase 1,SCO1)之间的相互作用,形成SCO1-LKB1-AMPK复合物,进而促进线粒体合成和脂肪酸氧化代谢.以上研究提示Cp与肝脏铜稳态密切相关,并且Cp可能是治疗NAFLD的潜在靶点.然而,关于Cp作为治疗NAFLD靶点的安全性与有效性仍需进一步验证. ...
... [29]还发现,抑制肝细胞中Cp的分泌可恢复肝脏铜含量至正常水平,并促进AMPK与细胞色素C氧化酶装配蛋白(synthesis of cytochrome C oxidase 1,SCO1)之间的相互作用,形成SCO1-LKB1-AMPK复合物,进而促进线粒体合成和脂肪酸氧化代谢.以上研究提示Cp与肝脏铜稳态密切相关,并且Cp可能是治疗NAFLD的潜在靶点.然而,关于Cp作为治疗NAFLD靶点的安全性与有效性仍需进一步验证. ...
1
... 由于Cp能携带肝脏的铜分布到机体的不同组织中,因此其在血中的水平升高可相应地引起肝脏中铜分布的不足.我们课题组之前的研究[29]发现,NAFLD患者血清中Cp水平上调导致肝细胞内的铜离子输出增多,从而引起肝脏铜含量降低.肝脏中铜含量的变化会引发一些疾病,如门克斯病(Menkes disease);该疾病是由ATP7A基因突变引起,会导致不同组织中铜的缺乏[30].此外,AIGNER等[31]的研究表明,NAFLD患者的肝脏铜浓度明显低于健康人群.HEFFERN等[32]使用Copper-Caged Luciferin-1(一种生物发光报告基因)进行体内组织特异性铜成像,监测活体动物体内的铜水平;他们也发现,在饮食诱导的NAFLD小鼠模型中,肝脏铜离子水平显著降低.TANG等[33]的研究表明,膳食铜离子缺乏会导致肝脂肪酸合成酶基因转录速度加快,从而提高肝脂质合成速度.核转录因子甾醇调节元件结合蛋白-1(sterol regulatory element-binding protein 1,SREBP-1)是脂肪酸合成的重要转录因子.在铜离子不足的情况下,成熟的SREBP-1会迁移到细胞核,从而促进脂肪酸从头合成的关键酶基因的转录. ...
1
... 由于Cp能携带肝脏的铜分布到机体的不同组织中,因此其在血中的水平升高可相应地引起肝脏中铜分布的不足.我们课题组之前的研究[29]发现,NAFLD患者血清中Cp水平上调导致肝细胞内的铜离子输出增多,从而引起肝脏铜含量降低.肝脏中铜含量的变化会引发一些疾病,如门克斯病(Menkes disease);该疾病是由ATP7A基因突变引起,会导致不同组织中铜的缺乏[30].此外,AIGNER等[31]的研究表明,NAFLD患者的肝脏铜浓度明显低于健康人群.HEFFERN等[32]使用Copper-Caged Luciferin-1(一种生物发光报告基因)进行体内组织特异性铜成像,监测活体动物体内的铜水平;他们也发现,在饮食诱导的NAFLD小鼠模型中,肝脏铜离子水平显著降低.TANG等[33]的研究表明,膳食铜离子缺乏会导致肝脂肪酸合成酶基因转录速度加快,从而提高肝脂质合成速度.核转录因子甾醇调节元件结合蛋白-1(sterol regulatory element-binding protein 1,SREBP-1)是脂肪酸合成的重要转录因子.在铜离子不足的情况下,成熟的SREBP-1会迁移到细胞核,从而促进脂肪酸从头合成的关键酶基因的转录. ...
1
... 由于Cp能携带肝脏的铜分布到机体的不同组织中,因此其在血中的水平升高可相应地引起肝脏中铜分布的不足.我们课题组之前的研究[29]发现,NAFLD患者血清中Cp水平上调导致肝细胞内的铜离子输出增多,从而引起肝脏铜含量降低.肝脏中铜含量的变化会引发一些疾病,如门克斯病(Menkes disease);该疾病是由ATP7A基因突变引起,会导致不同组织中铜的缺乏[30].此外,AIGNER等[31]的研究表明,NAFLD患者的肝脏铜浓度明显低于健康人群.HEFFERN等[32]使用Copper-Caged Luciferin-1(一种生物发光报告基因)进行体内组织特异性铜成像,监测活体动物体内的铜水平;他们也发现,在饮食诱导的NAFLD小鼠模型中,肝脏铜离子水平显著降低.TANG等[33]的研究表明,膳食铜离子缺乏会导致肝脂肪酸合成酶基因转录速度加快,从而提高肝脂质合成速度.核转录因子甾醇调节元件结合蛋白-1(sterol regulatory element-binding protein 1,SREBP-1)是脂肪酸合成的重要转录因子.在铜离子不足的情况下,成熟的SREBP-1会迁移到细胞核,从而促进脂肪酸从头合成的关键酶基因的转录. ...
1
... 由于Cp能携带肝脏的铜分布到机体的不同组织中,因此其在血中的水平升高可相应地引起肝脏中铜分布的不足.我们课题组之前的研究[29]发现,NAFLD患者血清中Cp水平上调导致肝细胞内的铜离子输出增多,从而引起肝脏铜含量降低.肝脏中铜含量的变化会引发一些疾病,如门克斯病(Menkes disease);该疾病是由ATP7A基因突变引起,会导致不同组织中铜的缺乏[30].此外,AIGNER等[31]的研究表明,NAFLD患者的肝脏铜浓度明显低于健康人群.HEFFERN等[32]使用Copper-Caged Luciferin-1(一种生物发光报告基因)进行体内组织特异性铜成像,监测活体动物体内的铜水平;他们也发现,在饮食诱导的NAFLD小鼠模型中,肝脏铜离子水平显著降低.TANG等[33]的研究表明,膳食铜离子缺乏会导致肝脂肪酸合成酶基因转录速度加快,从而提高肝脂质合成速度.核转录因子甾醇调节元件结合蛋白-1(sterol regulatory element-binding protein 1,SREBP-1)是脂肪酸合成的重要转录因子.在铜离子不足的情况下,成熟的SREBP-1会迁移到细胞核,从而促进脂肪酸从头合成的关键酶基因的转录. ...
1
... 我们课题组之前的研究[29]表明,Cp在调节肝脏TAG代谢中起到重要作用,并通过影响AMP依赖的蛋白激酶(adenosine 5′-monophosphate activated protein kinase,AMPK)信号通路发挥作用.AMPK是一种能量开关蛋白,调控细胞能量利用过程,包括脂质和葡萄糖代谢等.AMPK可激活过氧化物酶体增殖物活化受体γ共激活因子1α(peroxisome proliferator-activated receptor γ coactivator 1α,PGC1α),并通过与过氧化物酶体增殖物活化受体γ(peroxisome proliferator-activated receptor γ,PPARγ)或雌激素相关受体的相互作用,增加线粒体合成和促进脂肪酸的氧化.因此激活的AMPK可作为NAFLD的治疗靶点之一,通过活化AMPK在肝脏中发挥降脂作用[34-37].临床研究[38]发现,一种AMPK激活剂PXL770在口服给药后12周,可平均减少14.7%的肝脏脂肪含量.此外,我们课题组的研究[29]还发现,抑制肝细胞中Cp的分泌可恢复肝脏铜含量至正常水平,并促进AMPK与细胞色素C氧化酶装配蛋白(synthesis of cytochrome C oxidase 1,SCO1)之间的相互作用,形成SCO1-LKB1-AMPK复合物,进而促进线粒体合成和脂肪酸氧化代谢.以上研究提示Cp与肝脏铜稳态密切相关,并且Cp可能是治疗NAFLD的潜在靶点.然而,关于Cp作为治疗NAFLD靶点的安全性与有效性仍需进一步验证. ...
0
0
1
... 我们课题组之前的研究[29]表明,Cp在调节肝脏TAG代谢中起到重要作用,并通过影响AMP依赖的蛋白激酶(adenosine 5′-monophosphate activated protein kinase,AMPK)信号通路发挥作用.AMPK是一种能量开关蛋白,调控细胞能量利用过程,包括脂质和葡萄糖代谢等.AMPK可激活过氧化物酶体增殖物活化受体γ共激活因子1α(peroxisome proliferator-activated receptor γ coactivator 1α,PGC1α),并通过与过氧化物酶体增殖物活化受体γ(peroxisome proliferator-activated receptor γ,PPARγ)或雌激素相关受体的相互作用,增加线粒体合成和促进脂肪酸的氧化.因此激活的AMPK可作为NAFLD的治疗靶点之一,通过活化AMPK在肝脏中发挥降脂作用[34-37].临床研究[38]发现,一种AMPK激活剂PXL770在口服给药后12周,可平均减少14.7%的肝脏脂肪含量.此外,我们课题组的研究[29]还发现,抑制肝细胞中Cp的分泌可恢复肝脏铜含量至正常水平,并促进AMPK与细胞色素C氧化酶装配蛋白(synthesis of cytochrome C oxidase 1,SCO1)之间的相互作用,形成SCO1-LKB1-AMPK复合物,进而促进线粒体合成和脂肪酸氧化代谢.以上研究提示Cp与肝脏铜稳态密切相关,并且Cp可能是治疗NAFLD的潜在靶点.然而,关于Cp作为治疗NAFLD靶点的安全性与有效性仍需进一步验证. ...
2
... 我们课题组之前的研究[29]表明,Cp在调节肝脏TAG代谢中起到重要作用,并通过影响AMP依赖的蛋白激酶(adenosine 5′-monophosphate activated protein kinase,AMPK)信号通路发挥作用.AMPK是一种能量开关蛋白,调控细胞能量利用过程,包括脂质和葡萄糖代谢等.AMPK可激活过氧化物酶体增殖物活化受体γ共激活因子1α(peroxisome proliferator-activated receptor γ coactivator 1α,PGC1α),并通过与过氧化物酶体增殖物活化受体γ(peroxisome proliferator-activated receptor γ,PPARγ)或雌激素相关受体的相互作用,增加线粒体合成和促进脂肪酸的氧化.因此激活的AMPK可作为NAFLD的治疗靶点之一,通过活化AMPK在肝脏中发挥降脂作用[34-37].临床研究[38]发现,一种AMPK激活剂PXL770在口服给药后12周,可平均减少14.7%的肝脏脂肪含量.此外,我们课题组的研究[29]还发现,抑制肝细胞中Cp的分泌可恢复肝脏铜含量至正常水平,并促进AMPK与细胞色素C氧化酶装配蛋白(synthesis of cytochrome C oxidase 1,SCO1)之间的相互作用,形成SCO1-LKB1-AMPK复合物,进而促进线粒体合成和脂肪酸氧化代谢.以上研究提示Cp与肝脏铜稳态密切相关,并且Cp可能是治疗NAFLD的潜在靶点.然而,关于Cp作为治疗NAFLD靶点的安全性与有效性仍需进一步验证. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}