Objective ·To explore the activation methods and activity regulation mechanisms of the testisin zymogen in vitro, laying a foundation for further research on the physiological functions of testisin in organisms. Methods ·The eukaryotic expression plasmid for the mouse-derived testisin (mTN) zymogen was constructed through full-gene synthesis and subsequently transfected into eukaryotic HEK293S cells for expression. Purified recombinant testisin protein was obtained through methods such as nickel ion affinity chromatography. Site-directed mutagenesis was performed on the activation site and active center of the zymogen, and the activation mechanism of testisin was analyzed through enzymatic activity assays. Additionally, the activation rate of the protein was determined by altering the incubation conditions of the zymogen. The self-cleavage site of the protease was identified by using Edman degradation and site-directed mutagenesis. Finally, the regulatory mechanism of protease activity was examined by analyzing the proteolytic rate of the protein towards its substrates. Results ·The recombinant mTN zymogen was expressed in eukaryotic HEK293S cells, and high-purity, uniform recombinant protein was obtained through a two-step purification process. This protein was found to remain stable under acidic conditions but underwent self-activation in neutral or alkaline environments. The rate of activation was influenced by factors such as pH value and temperature. The self-activation process required on the integrity of the enzyme's activation site, Arg46, and its active center, Ser240. Additionally, the self-activation of the recombinant mTN zymogen was often accompanied by the self-cleavage at the 170/175 loop on the molecular surface, but this self-cleavage did not affect the activity of testisin. Furthermore, the optimal reaction pH for activated mTN was 8.0, with the protein remaining relatively stable near this pH. The optimal reaction temperature was 50 ℃, while the protein was best stored at temperatures below 30 ℃. Zn2+ and Ca2+ were found to significantly inhibit mTN activity. Conclusion ·An effective method for the preparation of recombinant mTN zymogen is successfully established, and it is discovered that the recombinant mTN undergoes self-activation and self-cleavage in vitro.
WAN Jiahui, ZHOU Aiwu. Mechanistic studies on regulation of the activity of GPI-anchored serine protease testisin. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2024, 44(12): 1504-1513 doi:10.3969/j.issn.1674-8115.2024.12.003
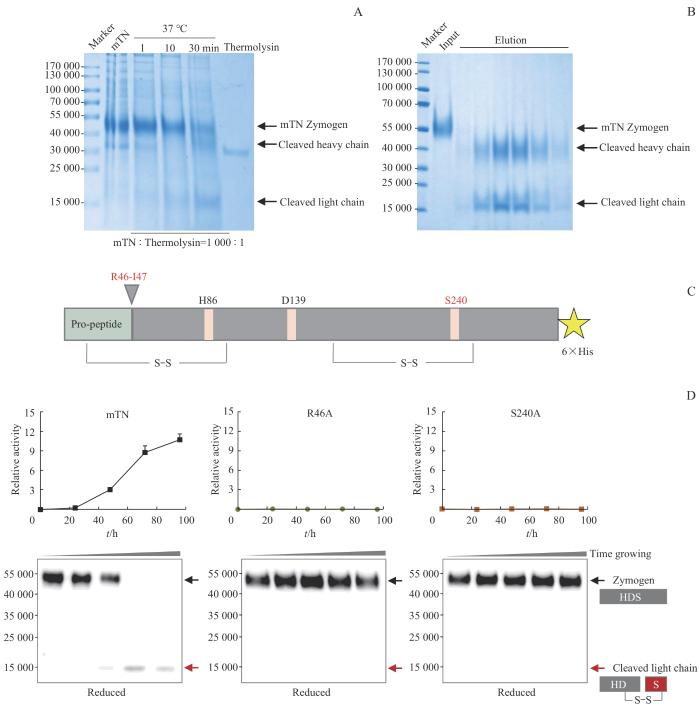
Note:A. SDS-PAGE of the preliminary activation tests of mTN under reducing conditions. B. SDS-PAGE of mTN obtained by Q anion exchange chromatography under reducing conditions. C.Schematic diagram of the molecular structure of mTN with two mutation sites prominently identified. D. Activation status of mTN and the two variants was detected every 24 h. The relative activity of mTN was derived from slope of the linear plot of absorbance at 405 nm vers incubation time during activity assay. Western blotting analysis revealed that a band of approximately 5 000 corresponded to the zymogen, while a band of approximately 1 500 corresponded to the degraded C-terminal fragment.
Fig 1
Self-activation verification of mTN in vitro
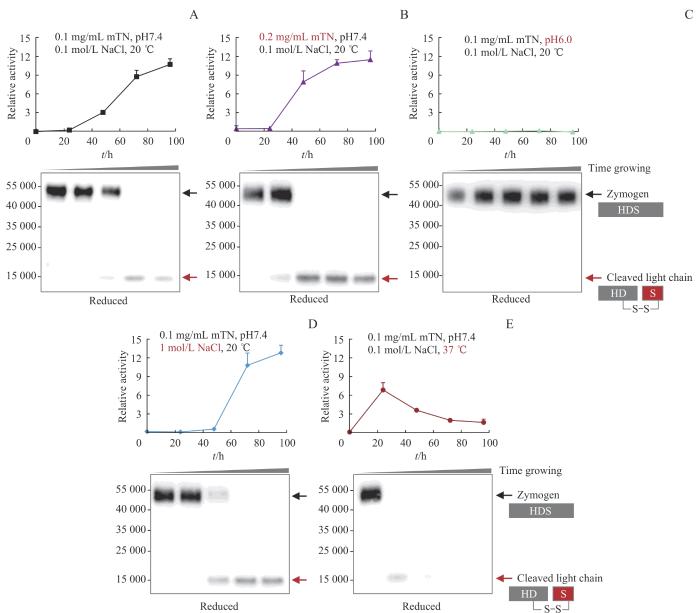
Note: Every 24 h, an equal amount of mTN under different incubation conditions was diluted into a substrate S2238 solution to detect the activation status of the protein. Western blotting was performed to assess protein cleavage under reducing conditions.
Fig 2
Influencing factors of mTN self-activation rate
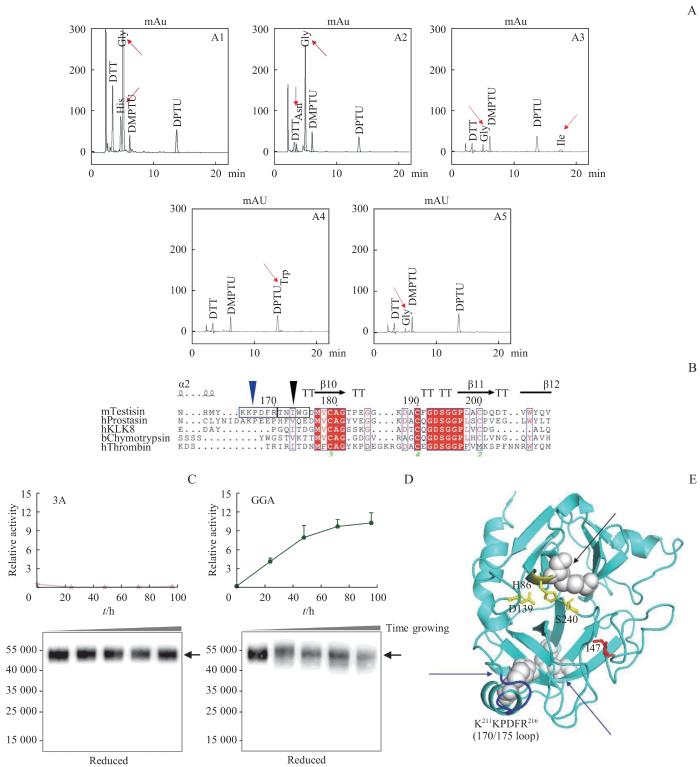
Note:A. The results for the N-terminal sequence of the light chain peptide of mTN (the first five amino acids). The red arrow indicates the measured amino acid signals. The Y-axis represents signal intensity, and the X-axis represents the testing time. Au—Absorbance unit.1 Au=1 000 mAu. B. The comparison result of the catalytic domain sequences of mTN with other serine proteases (partial). The five amino acids indicated by the black triangle corresponded to the first five amino acids of the N-terminal sequencing; the six amino acids indicated by the blue triangle corresponded to amino acids 211‒216 (KKPDFR) of the recombinant protein, where the cleavage site was thought to be located. C.The enzymatic activity trend and Western blotting result of the 3A mutant. D. The enzymatic activity trend and Western blotting result of the GGA mutant. E. Predicted structure of the mTN zymogen generated by Alphafold 2.
Fig 3
N-terminal sequencing results and validation of self-activation in mTN-3A & GGA mutants
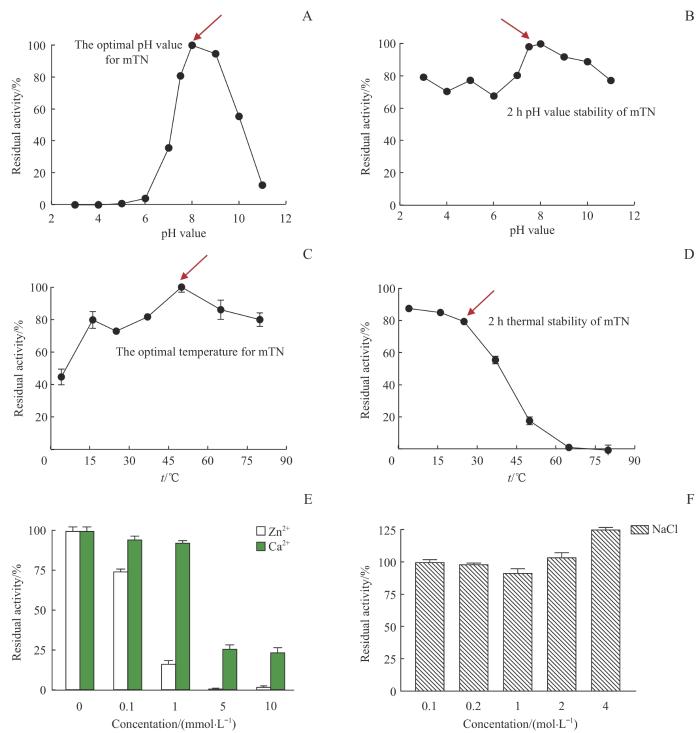
Note:A. The optimal pH value for mTN reaction. B. pH value stability of mTN. C. The optimal temperature for mTN reaction. D. Thermal stability of mTN. E. The effect of Zn2+ and Ca2+ on the activity of mTN. F. The effect of Na+ on the activity of mTN.
Fig 4
Effect of pH value, temperature and metal ions on mTN activity
ZHOU Aiwu supervised the research design and the revision of the paper. WAN Jiahui was responsible for the experimental work, data organisation, and writing and revising the paper. Both the authors have read the last version of paper and consented for submission.
利益冲突声明
两位作者声明不存在利益冲突。
COMPETING INTERESTS
Both authors disclose no relevant conflict of interests.
ANTALIS T M, BUZZA M S, HODGE K M, et al. The cutting edge: membrane-anchored serine protease activities in the pericellular microenvironment[J]. Biochem J, 2010, 428(3): 325-346.
VERGHESE G M, GUTKNECHT M F, CAUGHEY G H. Prostasin regulates epithelial monolayer function: cell-specific Gpld1-mediated secretion and functional role for GPI anchor[J]. Am J Physiol Cell Physiol, 2006, 291(6): C1258-C1270.
HONDA A, YAMAGATA K, SUGIURA S, et al. A mouse serine protease TESP5 is selectively included into lipid rafts of sperm membrane presumably as a glycosylphosphatidylinositol-anchored protein[J]. J Biol Chem, 2002, 277(19): 16976-16984.
HOOPER J D, NICOL D L, DICKINSON J L, et al. testisin, a new human serine proteinase expressed by premeiotic testicular germ cells and lost in testicular germ cell tumors[J]. Cancer Res, 1999, 59(13): 3199-3205.
TANG T, KMET M, CORRAL L, et al. testisin, a glycosyl-phosphatidylinositol-linked serine protease, promotes malignant transformation in vitro and in vivo[J]. Cancer Res, 2005, 65(3): 868-878.
YEOM S Y, JANG H L, LEE S J, et al. Interaction of testisin with maspin and its impact on invasion and cell death resistance of cervical cancer cells[J]. FEBS Lett, 2010, 584(8): 1469-1475.
AIMES R T, ZIJLSTRA A, HOOPER J D, et al. Endothelial cell serine proteases expressed during vascular morphogenesis and angiogenesis[J]. Thromb Haemost, 2003, 89(3): 561-572.
DRIESBAUGH K H, BUZZA M S, MARTIN E W, et al. Proteolytic activation of the protease-activated receptor (PAR)-2 by the glycosylphosphatidylinositol-anchored serine protease testisin[J]. J Biol Chem, 2015, 290(6): 3529-3541.
PEROUTKA R J, BUZZA M S, MUKHOPADHYAY S, et al. testisin/Prss21 deficiency causes increased vascular permeability and a hemorrhagic phenotype during luteal angiogenesis[J]. PLoS One, 2020, 15(6): e0234407.
DONG N Z, ZHOU T T, ZHANG Y, et al. Corin mutations K317E and S472G from preeclamptic patients alter zymogen activation and cell surface targeting.[Corrected][J]. J Biol Chem, 2014, 289(25): 17909-17916.
SHI Y E, TORRI J, YIEH L, et al. Identification and characterization of a novel matrix-degrading protease from hormone-dependent human breast cancer cells[J]. Cancer Res, 1993, 53(6): 1409-1415.
JIN X L, HIROSAKI T, LIN C Y, et al. Production of soluble matriptase by human cancer cell lines and cell surface activation of its zymogen by trypsin[J]. J Cell Biochem, 2005, 95(3): 632-647.
BENAUD C, DICKSON R B, LIN C Y. Regulation of the activity of matriptase on epithelial cell surfaces by a blood-derived factor[J]. Eur J Biochem, 2001, 268(5): 1439-1447.
AFAR D E, VIVANCO I, HUBERT R S, et al. Catalytic cleavage of the androgen-regulated TMPRSS2 protease results in its secretion by prostate and prostate cancer epithelia[J]. Cancer Res, 2001, 61(4): 1686-1692.
LUCAS J M, TRUE L, HAWLEY S, et al. The androgen-regulated type Ⅱ serine protease TMPRSS2 is differentially expressed and mislocalized in prostate adenocarcinoma[J]. J Pathol, 2008, 215(2): 118-125.
PAWAR N R, BUZZA M S, ANTALIS T M. Membrane-anchored serine proteases and protease-activated receptor-2-mediated signaling: co-conspirators in cancer progression[J]. Cancer Res, 2019, 79(2): 301-310.
TRUEBESTEIN L, TENNSTAEDT A, MÖNIG T, et al. Substrate-induced remodeling of the active site regulates human HTRA1 activity[J]. Nat Struct Mol Biol, 2011, 18(3): 386-388.
LECHTENBERG B C, MURRAY-RUST T A, JOHNSON D J, et al. Crystal structure of the prothrombinase complex from the venom of Pseudonaja textilis[J]. Blood, 2013, 122(16): 2777-2783.
REYDA S, SOHN C, KLEBE G, et al. Reconstructing the binding site of factor Ⅹa in trypsin reveals ligand-induced structural plasticity[J]. J Mol Biol, 2003, 325(5): 963-977.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}