Research progress in the role of β-arrestin 1 in the regulation of inflammatory response
YI Wenjing,, FAN Yixuan, QIU Jiatai, FU Xiaoyan, LIU Meifang,
Key Laboratory of Immune Microenvironment and Inflammatory Disease Research, School of Basic Medical Sciences, Shandong Second Medical University, Weifang 261053, China
The inflammatory response is a systemic process produced by the body in response to stimulation such as infection or injury. The inflammatory response is closely related to the pathogenesis of various diseases, and plays an important role in the progression and outcome of diseases. Under normal conditions, the inflammatory response can maintain tissue homeostasis when the body is not seriously damaged. However, an uncontrolled inflammatory response that occurs when the body is subjected to more severe stimuli may often cause serious harm to the body. The termination of the ideal inflammatory response occurs after the elimination of the inflammatory stimulus. Therefore, it is particularly important to effectively modulate the extent and scope of the inflammatory response. β-arrestin 1 (ARRB1) is a multifunctional regulatory protein that plays a key role in G protein-coupled receptor (GPCR) and non-GPCR-mediated signal transduction. Meanwhile, this protein also participates in the regulation of some immune cell development, differentiation and other functions. Although ARRB1 was originally thought to be a terminator of GPCR signal transduction and can inhibit the inflammatory response, recent studies suggest that its role in inflammatory responses is complex, with dual anti-inflammatory and pro-inflammatory effects. Based on this, this article reviews the relevant research on ARRB1 and the inflammatory response in recent years, focusing on the regulatory role of ARRB1 in signaling pathway transduction and immune cell development and differentiation function, as well as its mechanism of action in regulating the progression of inflammatory diseases, in order to provide new insights for the clinical precision treatment of inflammatory diseases and the screening of drug effect targets.
Keywords:β-arrestin 1 (ARRB1)
;
anti-inflammatory role
;
pro-inflammatory role
;
signaling pathway
;
immune cell
;
inflammatory factor
YI Wenjing, FAN Yixuan, QIU Jiatai, FU Xiaoyan, LIU Meifang. Research progress in the role of β-arrestin 1 in the regulation of inflammatory response. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2024, 44(12): 1593-1600 doi:10.3969/j.issn.1674-8115.2024.12.013
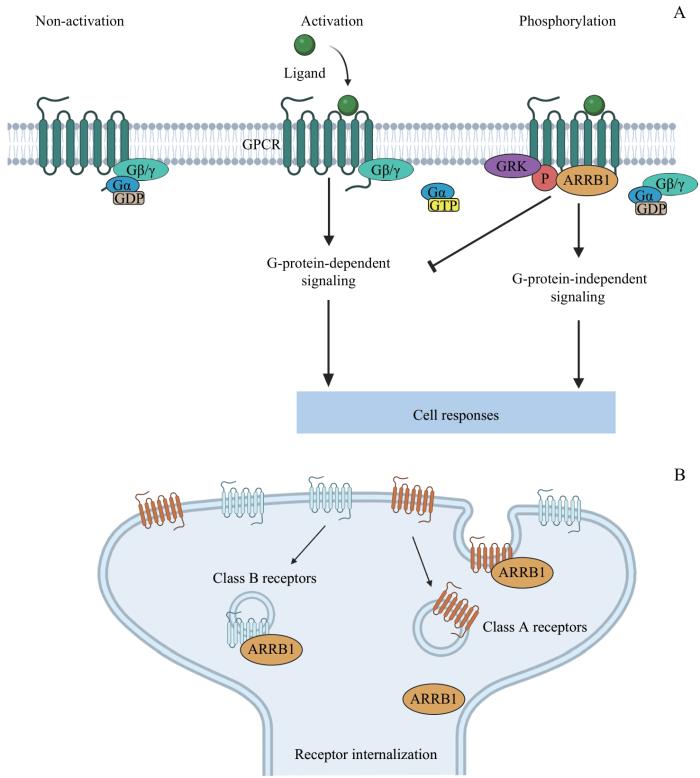
Note: A. ARRB1 is involved in GPCR desensitization and activation of G-protein-independent signaling pathways. B. ARRB1 is involved in receptor internalization.
Fig 1
Schematic diagram of the mechanism by which ARRBl regulates intracellular signal transduction
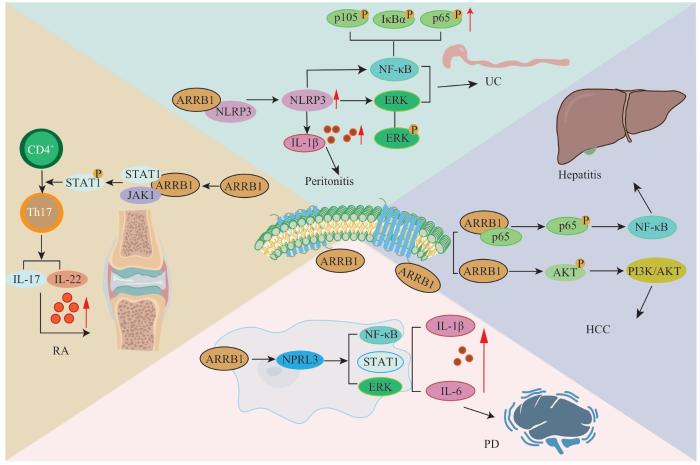
ARRB1促进炎症性疾病进展的作用机制(图3)可能涉及多种炎症相关信号通路(如NF-κB、STAT1等)的活化。近年来,在帕金森病小鼠模型中,研究人员发现ARRB1可直接上调胶质细胞中氮酶调节因子样蛋白3(nitrogen permease regulator like protein 3,NPRL3)的表达,促进下游NF-κB、STAT1信号通路的激活,进而增强胶质细胞介导的炎症反应程度[33]。有研究[34]在UC小鼠模型中发现,ARRB1可通过与核苷酸结合结构域富含亮氨酸重复序列和含热蛋白结构域受体3(nucleotide-binding domain leucine-rich repeat and pyrin domain-containing receptor 3,NLRP3)结合促进其寡聚化并使其合成增多,继而促进下游信号通路与促炎症因子的表达,最终导致结肠炎的恶化。同样,在小鼠腹膜炎模型的研究[35]中也发现,ARRB1可直接激活NLRP3并促进IL-1β的分泌,从而加重炎症反应。上述2项研究提示,NLRP3是ARRB1的重要靶蛋白,较高水平的ARRB1表达可加重由NLRP3介导的炎症反应。
YI Wenjing was responsible for the writing and revision of the manuscript. FAN Yixuan was responsible for the writing and literature combing. QIU Jiatai participated in the revision of the manuscript. LIU Meifang and FU Xiaoyan proposed the writing ideas. LIU Meifang was responsible for guiding the revision of the manuscript. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
CHEN S Z, HUANG M Y, ZHANG L M, et al. Inflammatory response signature score model for predicting immunotherapy response and pan-cancer prognosis[J]. Comput Struct Biotechnol J, 2024, 23: 369-383.
SCHROEDER H T, DE LEMOS MULLER C H, HECK T G, et al. Heat shock response during the resolution of inflammation and its progressive suppression in chronic-degenerative inflammatory diseases[J]. Cell Stress Chaperones, 2024, 29(1): 116-142.
FURMAN D, CAMPISI J, VERDIN E, et al. Chronic inflammation in the etiology of disease across the life span[J]. Nat Med, 2019, 25(12): 1822-1832.
YING W, FU W X, LEE Y S, et al. The role of macrophages in obesity-associated islet inflammation and β-cell abnormalities[J]. Nat Rev Endocrinol, 2020, 16(2): 81-90.
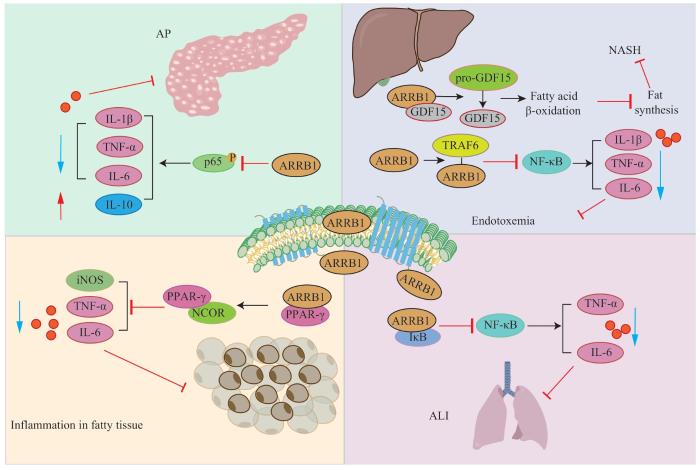
LI D J, TONG J, LI Y H, et al. Melatonin safeguards against fatty liver by antagonizing TRAFs-mediated ASK1 deubiquitination and stabilization in a β-arrestin-1 dependent manner[J]. J Pineal Res, 2019, 67(4): e12611.
DAWED A Y, MARI A, BROWN A, et al. Pharmacogenomics of GLP-1 receptor agonists: a genome-wide analysis of observational data and large randomised controlled trials[J]. Lancet Diabetes Endocrinol, 2023, 11(1): 33-41.
CHOI Y J, KIM J E, LEE S J, et al. Promotion of the inflammatory response in mid colon of complement component 3 knockout mice[J]. Sci Rep, 2022, 12(1): 1700.
MAFI A, KIM S K, GODDARD WA 3rd. The mechanism for ligand activation of the GPCR-G protein complex[J]. Proc Natl Acad Sci U S A, 2022, 119(18): e2110085119.
KO M J, CHIANG T, MUKADAM A A, et al. β-arrestin-dependent ERK signaling reduces anxiety-like and conditioned fear-related behaviors in mice[J]. Sci Signal, 2021, 14(694): eaba0245.
ZHANG Z, ZHONG X H, XIAO Y P, et al. MicroRNA-296 inhibits colorectal cancer cell growth and enhances apoptosis by targeting ARRB1-mediated AKT activation[J]. Oncol Rep, 2019, 41(1): 619-629.
LI J, WEI B, GUO A, et al. Deficiency of β-arrestin1 ameliorates collagen-induced arthritis with impaired TH17 cell differentiation[J]. Proc Natl Acad Sci U S A, 2013, 110(18): 7395-7400.
XU X, LEI Y M, CHEN L J, et al. Phosphorylation of NF-κBp65 drives inflammation-mediated hepatocellular carcinogenesis and is a novel therapeutic target[J]. J Exp Clin Cancer Res, 2021, 40(1): 253.
OAKLEY R H, LAPORTE S A, HOLT J A, et al. Differential affinities of visual arrestin, β arrestin1, and β arrestin2 for G protein-coupled receptors delineate two major classes of receptors[J]. J Biol Chem, 2000, 275(22): 17201-17210.
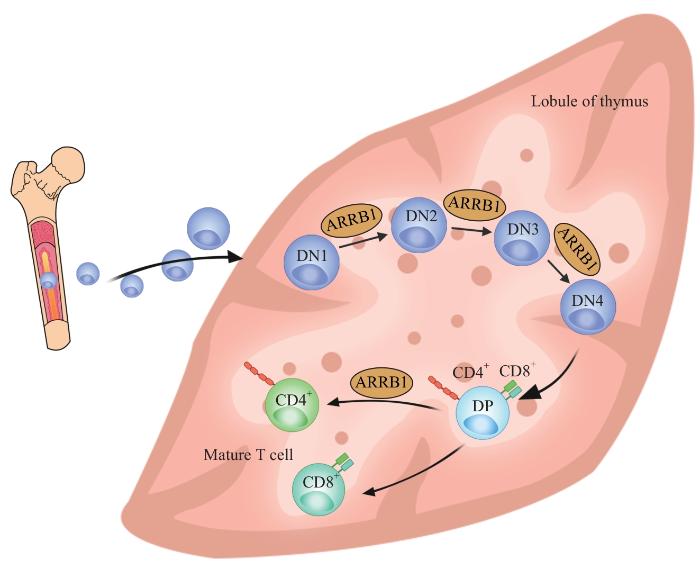
CHEN Y H, JIANG G J, GUO W, et al. Effects of Arrb1 knockout on the development of mice T lymphocytes[J]. Chinese Journal of Cell Biology, 2018, 40(3): 373-380.
GÓMEZ-MELERO S, GARCÍA-MACEIRA F I, GARCÍA-MACEIRA T, et al. Amino terminal recognition by a CCR6 chemokine receptor antibody blocks CCL20 signaling and IL-17 expression via β-arrestin[J]. BMC Biotechnol, 2021, 21(1): 41.
CAO Y P, HE W, LI X P, et al. Rosiglitazone protects against acetaminophen-induced acute liver injury by inhibiting multiple endoplasmic reticulum stress pathways[J]. Biomed Res Int, 2022, 2022: 6098592.
LEI Y M, WAN S Z, LIU H L, et al. ARRB1 suppresses the activation of hepatic macrophages via modulating endoplasmic reticulum stress in lipopolysaccharide-induced acute liver injury[J]. Cell Death Discov, 2021, 7(1): 223.
WU B S, ZHOU Q, HE Z Q, et al. Protective effect of the Abelmoschus manihot flower extract on DSS-induced ulcerative colitis in mice[J]. Evid Based Complement Alternat Med, 2021, 2021: 7422792.
FANG Y Q, JIANG Q L, LI S S, et al. Opposing functions of β-arrestin 1 and 2 in Parkinson's disease via microglia inflammation and Nprl3[J]. Cell Death Differ, 2021, 28(6): 1822-1836.
ZHANG Q, XU N, HU X, et al. Anti-colitic effects of Physalin B on dextran sodium sulfate-induced BALB/c mice by suppressing multiple inflammatory signaling pathways[J]. J Ethnopharmacol, 2020, 259: 112956.
YANG Y D, GUO Y W, TAN S W, et al. β-arrestin1 enhances hepatocellular carcinogenesis through inflammation-mediated Akt signalling[J]. Nat Commun, 2015, 6: 7369.
ZHANG Z C, XU X L, TIAN W F, et al. ARRB1 inhibits non-alcoholic steatohepatitis progression by promoting GDF15 maturation[J]. J Hepatol, 2020, 72(5): 976-989.
ZHUANG L N, HU W X, XIN S M, et al. β-arrestin-1 protein represses adipogenesis and inflammatory responses through its interaction with peroxisome proliferator-activated receptor-γ (PPARγ)[J]. J Biol Chem, 2011, 286(32): 28403-28413.
TAO L, LIN X Y, TAN S W, et al. β-arrestin1 alleviates acute pancreatitis via repression of NF-κBp65 activation[J]. J Gastroenterol Hepatol, 2019, 34(1): 284-292.
WITHEROW D S, GARRISON T R, MILLER W E, et al. β-arrestin inhibits NF-κB activity by means of its interaction with the NF-κB inhibitor IκBα[J]. Proc Natl Acad Sci U S A, 2004, 101(23): 8603-8607.
FAN H K, LUTTRELL L M, TEMPEL G E, et al. β-arrestins 1 and 2 differentially regulate LPS-induced signaling and pro-inflammatory gene expression[J]. Mol Immunol, 2007, 44(12): 3092-3099.
WANG M W, YANG Z, CHEN X, et al. Activation of PTH1R alleviates epididymitis and orchitis through Gq and β-arrestin-1 pathways[J]. Proc Natl Acad Sci U S A, 2021, 118(45): e2107363118.
SHU Y, WANG Y, LV W Q, et al. ARRB1-promoted NOTCH1 degradation is suppressed by oncomiR miR-223 in T-cell acute lymphoblastic leukemia[J]. Cancer Res, 2020, 80(5): 988-998.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}