目的·利用生物信息学方法探索骨关节炎与烟酰胺代谢相关基因之间的关系,找到具有诊断价值和治疗潜力的关键基因。方法·以“Osteoarthritis”为检索词,在GEO数据库中获取GSE12021、GSE55235和GSE55457数据集,将GSE55457作为验证集。去除GSE12021和GSE55235数据集的批次效应后,得到标准化的合并数据集,将其作为训练集,并在训练集中筛选出差异表达基因(differentially expressed genes,DEGs)。在GeneCards数据库和MSigDB数据库中获取所有烟酰胺代谢相关基因(nicotinamide metabolism-related genes,NMRGs)。将DEGs与NMRGs取交集,得到烟酰胺代谢相关差异表达基因(nicotinamide metabolism-related differentially expressed genes,NMRDEGs)。对训练集进行基因集富集分析(gene set enrichment analysis,GSEA),对NMRDEGs进行基因本体(Gene Ontology,GO)、京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)分析。通过LASSO(least absolute shrinkage and selection operator)和支持向量机(support vector machine,SVM)分析筛选出NMRDEGs关键基因,构建骨关节炎诊断模型,并用验证集GSE55457进行验证。通过单样本基因集富集分析(single sample gene set enrichment analysis,ssGSEA)分析免疫细胞的浸润类型。通过DGIdb数据库、ENCORI数据库和CHIPBase数据库对关键基因的mRNA进行相互作用网络和药物小分子预测。通过干扰小RNA(small interfering RNA,siRNA)敲降软骨细胞内NMRDEGs关键基因,用实时荧光定量聚合酶链反应(real-time fluorogenic quantitative polymerase chain reaction,RT-qPCR)检测关键基因敲降对软骨形成相关基因表达的影响。结果·发现了NAMPT、TIPARP等7个NMRDEGs。GO和KEGG分析富集到核因子κB信号通路和正向调节白细胞介素-1介导的信号通路等。GSEA富集到缺氧诱导因子-1转录因子通路(Hif1 Tfpathway)和多配体蛋白聚糖1(syndecan 1)通路等信号通路。LASSO分析和SVM分析共同筛选得到NPAS2、TIPARP和NAMPT关键基因并构建了骨关节炎诊断模型,验证集检验提示诊断模型诊断效果具有高准确度。ssGSEA免疫浸润分析的结果显示,巨噬细胞等15种免疫细胞存在显著差异(均P<0.05)。找到了7个针对关键基因的潜在药物小分子,19种与关键基因相互作用且上游基因与下游基因数量之和大于10的miRNA,19种与关键基因结合且上游基因与下游基因数量之和大于7的转录因子,27个聚类数>19的RNA结合蛋白。RT-qPCR结果显示,关键基因敲降会降低软骨形成相关基因的表达。结论·NPAS2、TIPARP和NAMPT为烟酰胺代谢相关的关键基因,可据此构建骨关节炎诊断模型。
关键词:骨关节炎
;
烟酰胺代谢
;
生物信息学
;
诊断模型
;
列线图
Abstract
Objective ·To explore the relationship between osteoarthritis and nicotinamide metabolism-related genes using bioinformatics analysis, and identify key genes with diagnostic value and therapeutic potential. Methods ·By using "Osteoarthritis" as a search term, GSE12021, GSE55235, and GSE55457 were obtained from the GEO database, with GSE55457 being used as the validation set. After removing batch effects from the GSE12021 and GSE55235 datasets, the standardized combined dataset was obtained and used as the training dataset. Differentially expressed genes (DEGs) were identified from the training dataset. All nicotinamide metabolism-related genes (NMRGs) were obtained from the GeneCards and MSigDB databases. The intersection of DEGs and NMRGs was taken to obtain nicotinamide metabolism-related differentially expressed genes (NMRDEGs). Gene set enrichment analysis (GSEA) was performed on the training dataset, while gene ontology (GO) and Kyoto encyclopedia of genes and genomes (KEGG) analysis were performed on NMRDEGs. Key genes were selected by using least absolute shrinkage and selection operator (LASSO) and support vector machine (SVM) analysis in NMRDEGs to build an osteoarthritis diagnosis model which was validated by using the GSE55457 dataset. Single sample gene set enrichment analysis (ssGSEA) was used to analyze the immune cell infiltration type. Interactions networks and drug molecule predictions were obtained for these key genes' mRNA with the DGIdb, ENCORI, and CHIPBase databases. siRNA was used to knock down the key genes in chondrocytes, and then real-time fluorescence quantitative polymerase chain reaction (RT-qPCR) was used to detect the expression of chondrogenesis-related genes. Results ·Seven NMRDEGs, including NAMPT, TIPARP, were discovered. GO and KEGG analysis enriched some signaling pathways, such as nuclear factor-κB signaling pathway and positive regulation of interleukin-1-mediated signaling pathway. GSEA enriched pathways such as Hif1 Tfpathway and syndecan 1 pathway. Key genes NPAS2, TIPARP, and NAMPT were identified through LASSO and SVM analysis, and used to construct an osteoarthritis diagnostic model. The validated results showed that the diagnostic model had high accuracy. Immune infiltration analysis results obtained by ssGSEA showed significant differences (all P<0.05) in 15 types of immune cells, including macrophages. Seven potential small molecules targeting key genes were identified, along with 19 miRNAs with the sum of upstream and downstream >10, 19 transcription factors with upstream and downstream >7, and 27 RNA binding proteins with clusterNum >19. The results of RT-qPCR showed that knocking down key genes reduced the expression of chondrogenesis-related genes. Conclusion ·Through bioinformatics analysis, key genes related to nicotinamide metabolism, NPAS2, TIPARP, and NAMPT, are discovered, and an osteoarthritis diagnostic model is constructed.
DENG Qingsong, ZHANG Changqing, TAO Shicong. Exploration of the relationship between nicotinamide metabolism-related genes and osteoarthritis. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2024, 44(2): 145-160 doi:10.3969/j.issn.1674-8115.2024.02.001
用R包clusterProfiler[16]对NMRDEGs进行基因本体(Gene Ontology,GO)和京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)注释分析,条目筛选标准为adj.P<0.1且错误发现率(false discovery rate,FDR;q值)<0.25,adj.P矫正方法为Benjamini-Hochberg。用Cytoscape对分析结果进行可视化展示。
1.4 基于对照分组的基因集富集分析
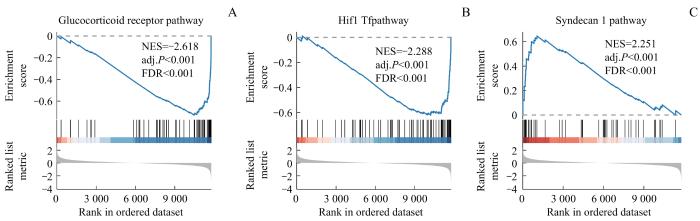
用R包clusterProfiler根据logFC值对训练集所有基因进行基因集富集分析(gene set enrichment analysis,GSEA)。GSEA分析中所使用的参数如下:种子为2023,计算次数为1 000,每个基因集最少含有的基因数为10,最多含有的基因数为500。在MSigDB数据库[17]中获取c2.all.v7.5.1.symbols.gmt基因集进行GSEA分析,筛选标准为adj.P<0.05且FDR值<0.25。
1.5 OA诊断模型的构建
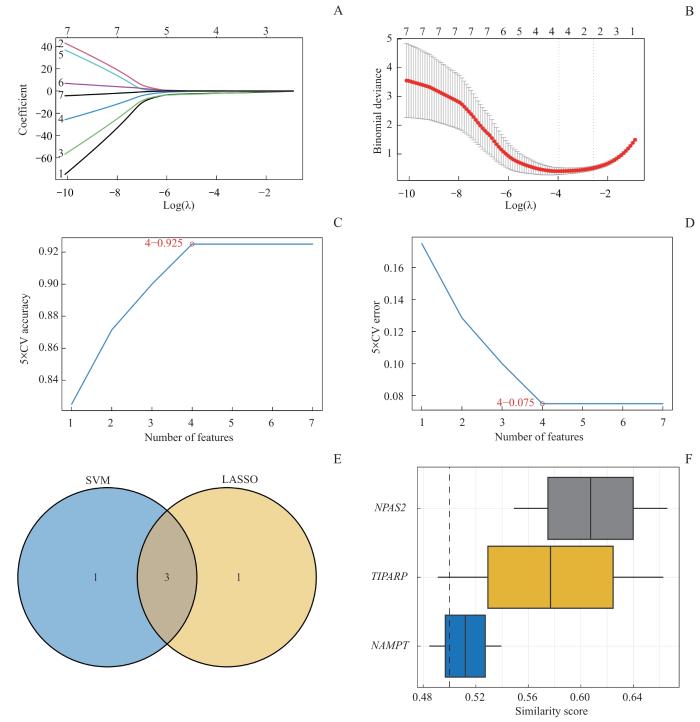
为得到NMRDEGs的诊断模型,用glmnet包[18]、参数family="binomial"和十折交叉验证执行LASSO(least absolute shrinkage and selection operator)回归[19],并运行1 000个周期以防止过度拟合。。通过支持向量机(support vector machine,SVM)算法构建SVM模型,并基于准确率最高和错误率最低的基因数量筛选NMRDEGs。之后再选取LASSO和SVM分析共同筛选得到的关键基因用于后续分析。
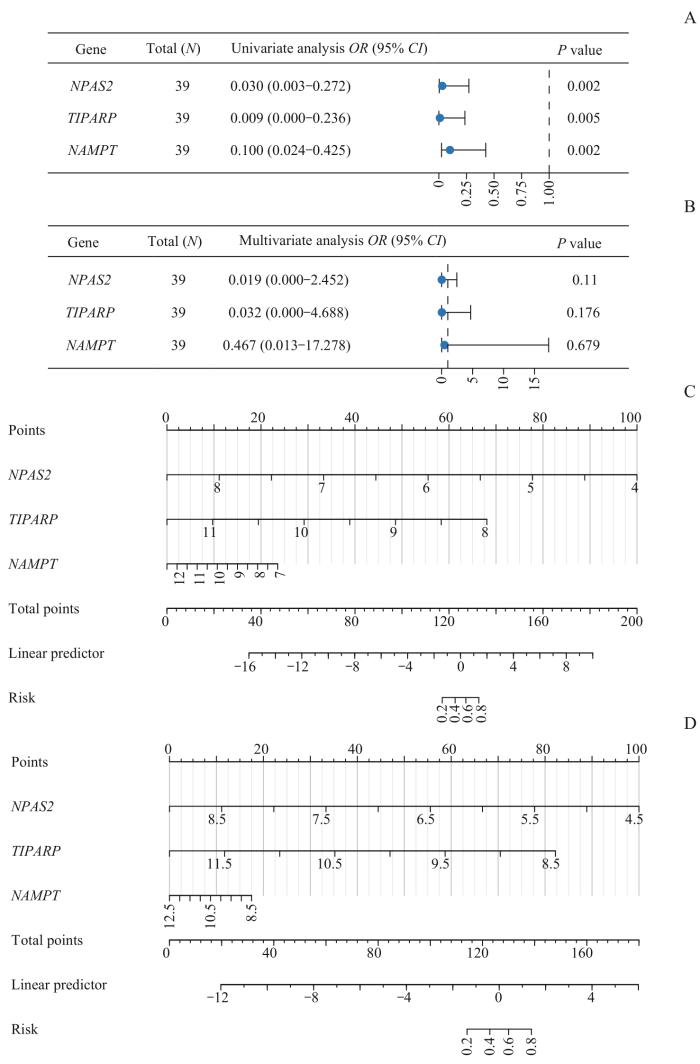
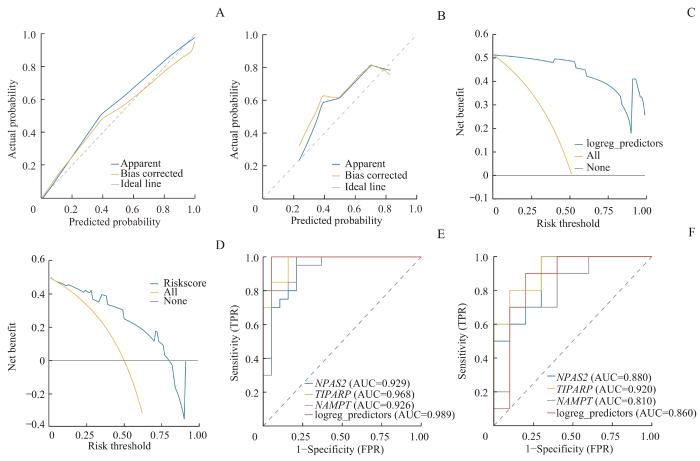
对关键基因进行Logistic回归分析并构建Logistic回归模型,根据Logistic回归分析结果用R包rms构建列线图[20]。用决策曲线分析(decision curve analysis,DCA)图评价Logistics回归模型的准确性和分辨力,并用R包ggDCA[21]绘制DCA图对Logistic回归模型的效果进行评估。用R包pROC绘制Logistic回归模型的受试者操作特征(receiver operating characteristic,ROC)曲线并计算ROC曲线下的面积(area under the curve,AUC)。最后,对关键基因进行功能相似性分析并绘制箱线图。
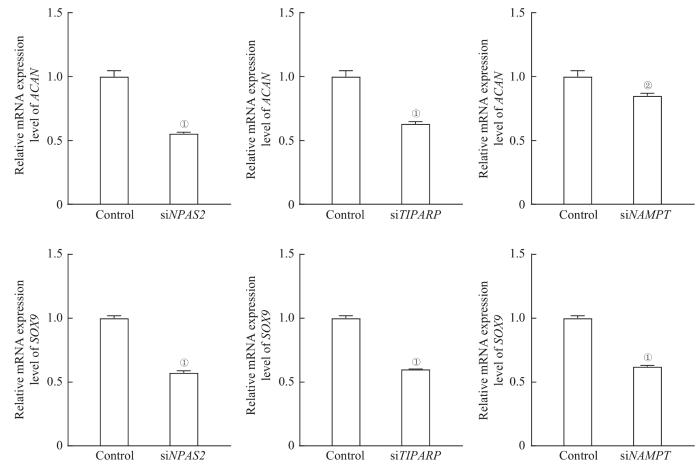
将人软骨细胞(C28/I2,德国默克)按照2×105个/孔的密度接种在6孔板中,按照EZ Trans RNA转染试剂(货号AC04L051,中国李记生物)的说明书分别转染针对关键基因的干扰小RNA(small interfering RNA,siRNA)(中国和元生物)。siRNA序列详见表1。用TRIzol(美国赛默飞)提取转染后软骨细胞的RNA,按照反转录试剂盒说明书(货号11141ES,中国翌圣生物)和实时荧光定量聚合酶链反应(real-time fluorogenic quantitative polymerase chain reaction,RT-qPCR)说明书(货号11204ES,中国翌圣生物)检测软骨形成相关基因聚集蛋白聚糖(aggrecan,ACAN)和Y染色体性别决定区-盒转录因子9(sex-determining region of Y chromosome-box transcription factor 9,SOX9)的表达。引物序列详见表2。
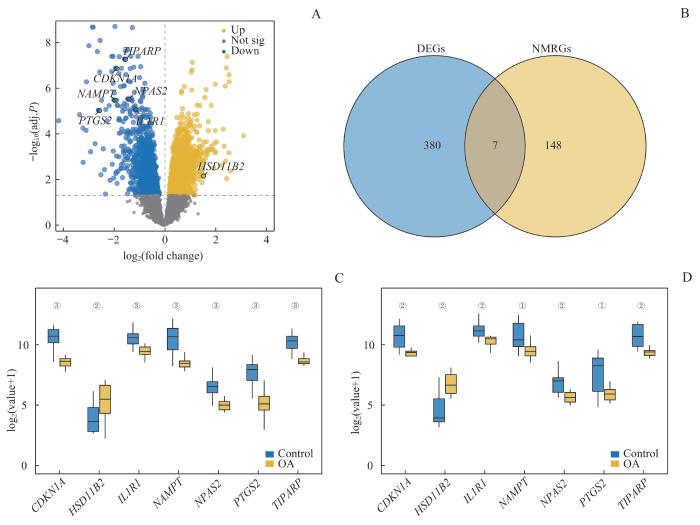
Note: A. Volcano plot of DEGs analysis in the OA group and the control group in the training dataset. B. Venn diagram of DEGs and NMRGs in the training dataset. C. Group comparison chart of NMRDEGs in the training dataset. D. Group comparison chart of NMRDEGs in the validation set GSE55457. ①P<0.05, ②P<0.01, ③P=0.000, comparison between the control group and the OA group.
Fig 2
DEGs expression analysis
2.2 GO和KEGG富集分析
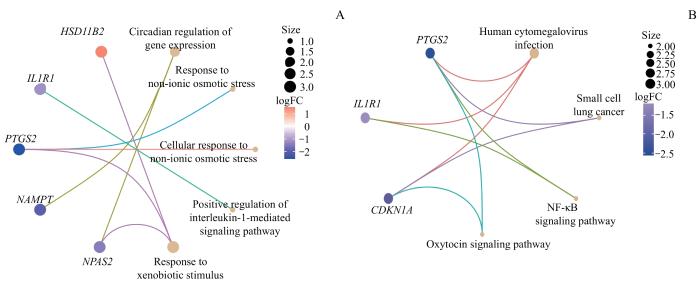
对7个NMRDEGs进行GO和KEGG富集分析,并将结果根据包含分子数量和logFC大小绘制成网络图(图3)。结果显示,7个NMRDEGs富集到了以下信号通路:正向调节白细胞介素-1介导的信号通路(positive regulation of interleukin-1-mediated signaling pathway)、对异种生物刺激的反应(response to xenobiotic stimulus)、基因表达的昼夜节律调控(circadian regulation of gene expression)、对非离子渗透压力的反应(response to non-ionic osmotic stress)、细胞对非离子渗透压力的反应(cellular response to non-ionic osmotic stress)、核因子κB信号通路(nuclear factor-κB signaling pathway,NF-κB signaling pathway)、人类巨细胞病毒感染(human cytomegalovirus infection)、小细胞肺癌(small cell lung cancer)、催产素信号通路(oxytocin signaling pathway)。
Note: A. The results of GO enrichment analysis combined with difference analysis results of NMRDEGs logFC network diagram display. B. The pathway KEGG enrichment analysis results of NMRDEGs combined with the difference analysis results of logFC network diagram display.
Note: A. LASSO regressor trajectories for NMRDEGs. B. LASSO regression diagnostic model diagram. C. The number of genes with the highest accuracy rate obtained by the SVM algorithm. D. The number of genes with the lowest error rate obtained by the SVM algorithm. E. Venn diagram of the intersection of SVM and LASSO models. F. Boxplot of functional similarity analysis of key genes.
Note: A. The key genes are included in the forest plot of the single factor Logistic regression model. B. The key genes are included in the forest plot of the multi-factor Logistic regression model. C. The nomogram of the key genes and Logistic predictive scoring model in the training dataset. D. The nomogram of the key genes and Logistic predictive value scoring model in the validation set GSE55457.
Note: A. Calibration plot of the Logistic model in the training dataset. B. Calibration plot of the Logistic model in the validation set GSE55457. C. DCA plot of the Logistic model in the training dataset. D. DCA plot of the Logistic model in the validation set GSE55457. E. The ROC curve of the Logistic model in the training dataset. F. The ROC curve of the Logistic model in the validation set GSE55457. FPR—false positive rate.
Fig 7
Validation analysis of OA diagnostic model
2.5 基于诊断模型得分分组的GSEA分析
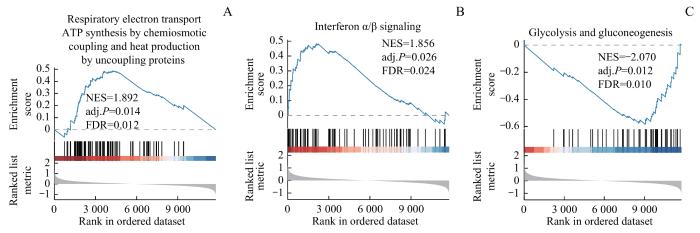
为了确定训练集高风险组和低风险组的Logistic诊断模型差异对OA样本中所有基因和生物过程表达的影响,通过GSEA分析了训练集中OA样本的基因表达和生物过程的关系。结果显示,高风险组和低风险组中的基因在化学渗透偶联的呼吸电子传递ATP合成和通过解偶联蛋白产生热量(respiratory electron transport ATP synthesis by chemiosmotic coupling and heat production by uncoupling proteins)(图8A)、干扰素α/β信号转导(interferon α/β signaling)(图8B)、糖酵解和糖原异生(glycolysis and gluconeogenesis)(图8C)等通路中显著富集。
Note: A. Respiratory electron transport ATP synthesis by chemiosmotic coupling and heat production by uncoupling proteins. B. Interferon α/β signaling. C. Glycolysis and gluconeogenesis.
Fig 8
GSEA based on high-low risk stratification
2.6 ssGSEA免疫浸润分析
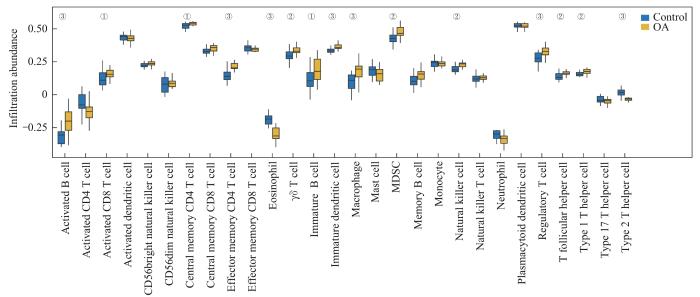
为了探究OA组和对照组在训练集中免疫浸润的差异,使用ssGSEA算法计算对照组和OA组样本中28种免疫细胞浸润丰度,然后用Mann-Whitney U检验分析其在不同组中的丰度差异。图9表明,在训练集中,15种免疫细胞类型的浸润丰度存在显著差异(均P<0.05),分别为2型辅助性T细胞(type 2 T cells,Th2细胞)、Th1细胞、滤泡辅助T细胞、调节性T细胞(regulatory T cells,Tregs)、自然杀伤细胞(natural killer cells,NK细胞)、髓源性抑制细胞(myeloid-derived suppressor cells,MDSCs)、巨噬细胞、未成熟树突状细胞、未成熟B细胞、γδT细胞、嗜酸性粒细胞、效应记忆CD4T细胞、中央记忆CD4T细胞、活化CD8T细胞和活化B细胞。
Note: Group comparison chart showing the differences in immune infiltration between the OA group and the control group in the training dataset. ①P<0.05, ②P<0.01, ③P=0.000.
Fig 9
Training dataset immune infiltration analysis by ssGSEA algorithm
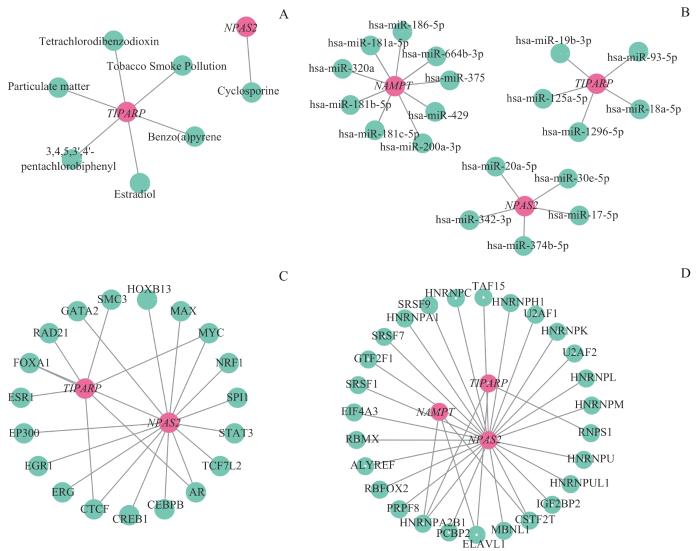
Note: A. mRNA-drug interaction network between key genes and small molecules. B. mRNA-miRNA interaction network of key genes and miRNA. C. mRNA-TF interaction network of key genes and transcription factors. D. The mRNA-RBP interaction network of key genes and RBP.
Fig 10
mRNA interactome network and drug prediction
除了NAMPT,关键基因NPAS2和TIPARP在OA中发挥的作用也同样重要。神经元PAS结构域蛋白2(neuronal PAS domain protein 2,NPAS2)是一种调节哺乳动物昼夜节律的转录因子,能够通过与脑和肌肉ARNT样蛋白1(brain and muscle arnt-like protein 1,BMAL1)形成二聚体参与昼夜节律的调节[29]。研究表明,以NAD(H)和NADP(H)为代表的细胞还原氧化状态调节CLOCK/BMAL1和NPAS2/BMAL1的转录活性,NAD还原形式能够提高NPAS2的DNA结合活性,而氧化形式降低了NPAS2的DNA结合活性[30]。TCCD诱导的多聚腺苷二磷酸核糖聚合酶(TCDD inducible poly-ADP-ribose polymerase,TIPARP)是一种单ADP-核糖基转移酶,是PARP家族成员,受芳烃受体(aryl hydrocarbon receptor,AHR)调控,其详细功能及其在调节转录中的作用尚不清楚。有研究表明,TIPARP是HIF-1的靶标,充当缺氧信号转导的负反馈调节剂,能够以ADP-核糖基化依赖性方式形成球形核体并募集HIF-1α和E3泛素连接酶HUWE1,导致HIF-1α的泛素化和降解[31]。此外研究[32]发现,OA中的TIPARP表达显著降低。
在本研究筛选出的另外4个NMRDEGs中,IL-1受体1型(interleukin 1 receptor type 1,IL1R1)是炎症因子IL-1β发挥作用的重要受体,在OA的炎症进展和疼痛中发挥重要作用[33]。11-β羟基类固醇脱氢酶2(11-β hydroxysteroid dehydrogenase 2,HSD11B2)是一种负责将皮质醇转化为非活性形式可的松的微粒体酶复合物,能够调节皮质醇水平。前列腺素内过氧化物合酶2(prostaglandin-endoperoxide synthase 2,PTGS2),又称环氧酶2(cyclooxygenase-2,COX-2),参与前列腺素的形成。在OA中,PTGS2、MMP1、MMP3、MMP9等一些分解代谢因子在HIF-2α的调节下介导了软骨破坏[34]。同时PTGS2的表达也受到NADPH的调控,NADPH能够通过抑制上游基因NF-κB,抑制PTGS2的表达[35]。CDKN1A又名p21,是细胞周期蛋白依赖性激酶抑制剂,能够阻滞细胞周期,同时细胞内p21常作为细胞衰老的生物标志物。在OA患者的软骨细胞中p21的转录和蛋白表达水平显著提高,进而诱导衰老软骨细胞分泌大量的衰老相关分泌表型,进一步加重OA[36]。
DENG Qingsong was responsible for data analysis and paper writing. ZHANG Changqing was responsible for paper review. TAO Shicong was responsible for experimental design and paper editing. All authors have read the final manuscript and agreed to the submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
REVOLLO J R, GRIMM A A, IMAI S. The NAD biosynthesis pathway mediated by nicotinamide phosphoribosyltransferase regulates Sir2 activity in mammalian cells[J]. J Biol Chem, 2004, 279(49): 50754-50763.
GARTEN A, SCHUSTER S, PENKE M, et al. Physiological and pathophysiological roles of NAMPT and NAD metabolism[J]. Nat Rev Endocrinol, 2015, 11(9): 535-546.
YANG S, RYU J H, OH H, et al. NAMPT (visfatin), a direct target of hypoxia-inducible factor-2α, is an essential catabolic regulator of osteoarthritis[J]. Ann Rheum Dis, 2015, 74(3): 595-602.
PRESLE N, POTTIE P, DUMOND H, et al. Differential distribution of adipokines between serum and synovial fluid in patients with osteoarthritis. Contribution of joint tissues to their articular production[J]. Osteoarthritis Cartilage, 2006, 14(7): 690-695.
HUBER R, HUMMERT C, GAUSMANN U, et al. Identification of intra-group, inter-individual, and gene-specific variances in mRNA expression profiles in the rheumatoid arthritis synovial membrane[J]. Arthritis Res Ther, 2008, 10(4): R98.
WOETZEL D, HUBER R, KUPFER P, et al. Identification of rheumatoid arthritis and osteoarthritis patients by transcriptome-based rule set generation[J]. Arthritis Res Ther, 2014, 16(2): R84.
STELZER G, ROSEN N, PLASCHKES I, et al. The GeneCards suite: from gene data mining to disease genome sequence analyses[J]. Curr Protoc Bioinformatics, 2016, 54: 1.30.1-1.30.33.
LEEK J T, JOHNSON W E, PARKER H S, et al. The sva package for removing batch effects and other unwanted variation in high-throughput experiments[J]. Bioinformatics, 2012, 28(6): 882-883.
RITCHIE M E, PHIPSON B, WU D, et al. Limma powers differential expression analyses for RNA-sequencing and microarray studies[J]. Nucleic Acids Res, 2015, 43(7): e47.
YANG L, QU Q, HAO Z, et al. Powerful identification of large quantitative trait loci using genome-wide R/glmnet-based regression[J]. J Hered, 2022, 113(4): 472-478.
NARALA S, LI S Q, KLIMAS N K, et al. Application of least absolute shrinkage and selection operator logistic regression for the histopathological comparison of chondrodermatitis nodularis helicis and hyperplastic actinic keratosis[J]. J Cutan Pathol, 2021, 48(6): 739-744.
SUN L, PANG Y, WANG X, et al. Ablation of gut microbiota alleviates obesity-induced hepatic steatosis and glucose intolerance by modulating bile acid metabolism in hamsters[J]. Acta Pharm Sin B, 2019, 9(4): 702-710.
XIAO B, LIU L, LI A, et al. Identification and verification of immune-related gene prognostic signature based on ssGSEA for osteosarcoma[J]. Front Oncol, 2020, 10: 607622.
FRESHOUR S L, KIWALA S, COTTO K C, et al. Integration of the Drug-Gene Interaction Database (DGIdb 4.0) with open crowdsource efforts[J]. Nucleic Acids Res, 2021, 49(D1): D1144-D1151.
ZHOU K R, LIU S, SUN W J, et al. ChIPBase v2.0: decoding transcriptional regulatory networks of non-coding RNAs and protein-coding genes from ChIP-seq data[J]. Nucleic Acids Res, 2017, 45(D1): D43-D50.
WANG Q H, LI Y, DOU D Y, et al. Nicotinamide mononucleotide-elicited NAMPT signaling activation aggravated adjuvant-induced arthritis in rats by affecting peripheral immune cells differentiation[J]. Int Immunopharmacol, 2021, 98: 107856.
NOWELL M, EVANS L, WILLIAMS A. PBEF/NAMPT/visfatin: a promising drug target for treating rheumatoid arthritis?[J]. Future Med Chem, 2012, 4(6): 751-769.
BECKER-KRAIL D D, PAREKH P K, KETCHESIN K D, et al. Circadian transcription factor NPAS2 and the NAD+-dependent deacetylase SIRT1 interact in the mouse nucleus accumbens and regulate reward[J]. Eur J Neurosci, 2022, 55(3): 675-693.
ZHANG L, CAO J, DONG L, et al. TiPARP forms nuclear condensates to degrade HIF-1α and suppress tumorigenesis[J]. Proc Natl Acad Sci USA, 2020, 117(24): 13447-13456.
SWAHN H, OLMER M, LOTZ M K. RNA-binding proteins that are highly expressed and enriched in healthy cartilage but suppressed in osteoarthritis[J]. Front Cell Dev Biol, 2023, 11: 1208315.
WYATT L A, NWOSU L N, WILSON D, et al. Molecular expression patterns in the synovium and their association with advanced symptomatic knee osteoarthritis[J]. Osteoarthritis Cartilage, 2019, 27(4): 667-675.
YANG S, KIM J, RYU J H, et al. Hypoxia-inducible factor-2α is a catabolic regulator of osteoarthritic cartilage destruction[J]. Nat Med, 2010, 16(6): 687-693.
QIN Y Y, LI M, FENG X, et al. Combined NADPH and the NOX inhibitor apocynin provides greater anti-inflammatory and neuroprotective effects in a mouse model of stroke[J]. Free Radic Biol Med, 2017, 104: 333-345.
CORYELL P R, DIEKMAN B O, LOESER R F. Mechanisms and therapeutic implications of cellular senescence in osteoarthritis[J]. Nat Rev Rheumatol, 2021, 17(1): 47-57.
SACTA M A, THARMALINGAM B, COPPO M, et al. Gene-specific mechanisms direct glucocorticoid-receptor-driven repression of inflammatory response genes in macrophages[J]. Elife, 2018, 7: e34864.
DIBATTISTA J A, MARTEL-PELLETIER J, ANTAKLY T, et al. Reduced expression of glucocorticoid receptor levels in human osteoarthritic chondrocytes. Role in the suppression of metalloprotease synthesis[J]. J Clin Endocrinol Metab, 1993, 76(5): 1128-1134.
PELLETIER J P, DIBATTISTA J A, RANGER P, et al. Modulation of the expression of glucocorticoid receptors in synovial fibroblasts and chondrocytes by prostaglandins and NSAIDs[J]. Am J Ther, 1996, 3(2): 115-119.
OKADA K, MORI D, MAKII Y, et al. Hypoxia-inducible factor-1 α maintains mouse articular cartilage through suppression of NF-κB signaling[J]. Sci Rep, 2020, 10(1): 5425.
ZHANG H, WANG L, CUI J, et al. Maintaining hypoxia environment of subchondral bone alleviates osteoarthritis progression[J]. Sci Adv, 2023, 9(14): eabo7868.
PATTERSON A M, CARTWRIGHT A, DAVID G, et al. Differential expression of syndecans and glypicans in chronically inflamed synovium[J]. Ann Rheum Dis, 2008, 67(5): 592-601.
KAUFMAN J, CARIC D, VUKOJEVIC K. Expression pattern of Syndecan-1 and HSP-70 in hip tissue of patients with osteoarthritis[J]. J Orthop, 2019, 17: 134-138.
SALMINEN-MANKONEN H, SÄÄMÄNEN A M, JALKANEN M, et al. Syndecan-1 expression is upregulated in degenerating articular cartilage in a transgenic mouse model for osteoarthritis[J]. Scand J Rheumatol, 2005, 34(6): 469-474.
ZHANG Y Q, YANG Y, WANG C Z, et al. Identification of diagnostic biomarkers of osteoarthritis based on multi-chip integrated analysis and machine learning[J]. DNA Cell Biol, 2020, 39(12): 2245-2256.
HAN Y, WU J, GONG Z, et al. Identification and development of a novel 5-gene diagnostic model based on immune infiltration analysis of osteoarthritis[J]. J Transl Med, 2021, 19(1): 522.
JAIME P, GARCÍA-GUERRERO N, ESTELLA R, et al. CD56+/CD16- natural killer cells expressing the inflammatory protease granzyme A are enriched in synovial fluid from patients with osteoarthritis[J]. Osteoarthritis Cartilage, 2017, 25(10): 1708-1718.
WANG H, ZENG Y, ZHANG M, et al. CD56brightCD16- natural killer cells are shifted toward an IFN-γ-promoting phenotype with reduced regulatory capacity in osteoarthritis[J]. Hum Immunol, 2019, 80(10): 871-877.
MORADI B, SCHNATZER P, HAGMANN S, et al. CD4+ CD25⁺/highCD127low/⁻ regulatory T cells are enriched in rheumatoid arthritis and osteoarthritis joints: analysis of frequency and phenotype in synovial membrane, synovial fluid and peripheral blood[J]. Arthritis Res Ther, 2014, 16(2): R97.
ZHANG L, LI Y G, LI Y H, et al. Increased frequencies of Th22 cells as well as Th17 cells in the peripheral blood of patients with ankylosing spondylitis and rheumatoid arthritis[J]. PLoS One, 2012, 7(4): e31000.
DOLHAIN R J E M, van der HEIDEN A N, TER HAAR N T, et al. Shift toward T lymphocytes with a T helper 1 cytokine-secretion profile in the joints of patients with rheumatoid arthritis[J]. Arthritis Rheum, 1996, 39(12): 1961-1969.
NEES T A, ZHANG J A, PLATZER H, et al. Infiltration profile of regulatory T cells in osteoarthritis-related pain and disability[J]. Biomedicines, 2022, 10(9): 2111.
BAGRI N K. Cyclosporine for systemic onset juvenile idiopathic arthritis: current stand and future directions[J]. Indian J Pediatr, 2019, 86(7): 576-577.
MANEIX L, BEAUCHEF G, SERVENT A, et al. 17β-oestradiol up-regulates the expression of a functional UDP-glucose dehydrogenase in articular chondrocytes: comparison with effects of cytokines and growth factors[J]. Rheumatology (Oxford), 2008, 47(3): 281-288.
CLAASSEN H, SCHÜNKE M, KURZ B. Estradiol protects cultured articular chondrocytes from oxygen-radical-induced damage[J]. Cell Tissue Res, 2005, 319(3): 439-445.
PANG H W, CHEN S H, KLYNE D M, et al. Low back pain and osteoarthritis pain: a perspective of estrogen[J]. Bone Res, 2023, 11(1): 42.
... 对关键基因进行Logistic回归分析并构建Logistic回归模型,根据Logistic回归分析结果用R包rms构建列线图[20].用决策曲线分析(decision curve analysis,DCA)图评价Logistics回归模型的准确性和分辨力,并用R包ggDCA[21]绘制DCA图对Logistic回归模型的效果进行评估.用R包pROC绘制Logistic回归模型的受试者操作特征(receiver operating characteristic,ROC)曲线并计算ROC曲线下的面积(area under the curve,AUC).最后,对关键基因进行功能相似性分析并绘制箱线图. ...
1
... 对关键基因进行Logistic回归分析并构建Logistic回归模型,根据Logistic回归分析结果用R包rms构建列线图[20].用决策曲线分析(decision curve analysis,DCA)图评价Logistics回归模型的准确性和分辨力,并用R包ggDCA[21]绘制DCA图对Logistic回归模型的效果进行评估.用R包pROC绘制Logistic回归模型的受试者操作特征(receiver operating characteristic,ROC)曲线并计算ROC曲线下的面积(area under the curve,AUC).最后,对关键基因进行功能相似性分析并绘制箱线图. ...
1
... 用单样本基因集富集分析(single sample gene set enrichment analysis,ssGSEA)算法[22]量化28种免疫细胞在训练集中浸润的相对丰度. ...
... 除了NAMPT,关键基因NPAS2和TIPARP在OA中发挥的作用也同样重要.神经元PAS结构域蛋白2(neuronal PAS domain protein 2,NPAS2)是一种调节哺乳动物昼夜节律的转录因子,能够通过与脑和肌肉ARNT样蛋白1(brain and muscle arnt-like protein 1,BMAL1)形成二聚体参与昼夜节律的调节[29].研究表明,以NAD(H)和NADP(H)为代表的细胞还原氧化状态调节CLOCK/BMAL1和NPAS2/BMAL1的转录活性,NAD还原形式能够提高NPAS2的DNA结合活性,而氧化形式降低了NPAS2的DNA结合活性[30].TCCD诱导的多聚腺苷二磷酸核糖聚合酶(TCDD inducible poly-ADP-ribose polymerase,TIPARP)是一种单ADP-核糖基转移酶,是PARP家族成员,受芳烃受体(aryl hydrocarbon receptor,AHR)调控,其详细功能及其在调节转录中的作用尚不清楚.有研究表明,TIPARP是HIF-1的靶标,充当缺氧信号转导的负反馈调节剂,能够以ADP-核糖基化依赖性方式形成球形核体并募集HIF-1α和E3泛素连接酶HUWE1,导致HIF-1α的泛素化和降解[31].此外研究[32]发现,OA中的TIPARP表达显著降低. ...
1
... 除了NAMPT,关键基因NPAS2和TIPARP在OA中发挥的作用也同样重要.神经元PAS结构域蛋白2(neuronal PAS domain protein 2,NPAS2)是一种调节哺乳动物昼夜节律的转录因子,能够通过与脑和肌肉ARNT样蛋白1(brain and muscle arnt-like protein 1,BMAL1)形成二聚体参与昼夜节律的调节[29].研究表明,以NAD(H)和NADP(H)为代表的细胞还原氧化状态调节CLOCK/BMAL1和NPAS2/BMAL1的转录活性,NAD还原形式能够提高NPAS2的DNA结合活性,而氧化形式降低了NPAS2的DNA结合活性[30].TCCD诱导的多聚腺苷二磷酸核糖聚合酶(TCDD inducible poly-ADP-ribose polymerase,TIPARP)是一种单ADP-核糖基转移酶,是PARP家族成员,受芳烃受体(aryl hydrocarbon receptor,AHR)调控,其详细功能及其在调节转录中的作用尚不清楚.有研究表明,TIPARP是HIF-1的靶标,充当缺氧信号转导的负反馈调节剂,能够以ADP-核糖基化依赖性方式形成球形核体并募集HIF-1α和E3泛素连接酶HUWE1,导致HIF-1α的泛素化和降解[31].此外研究[32]发现,OA中的TIPARP表达显著降低. ...
1
... 除了NAMPT,关键基因NPAS2和TIPARP在OA中发挥的作用也同样重要.神经元PAS结构域蛋白2(neuronal PAS domain protein 2,NPAS2)是一种调节哺乳动物昼夜节律的转录因子,能够通过与脑和肌肉ARNT样蛋白1(brain and muscle arnt-like protein 1,BMAL1)形成二聚体参与昼夜节律的调节[29].研究表明,以NAD(H)和NADP(H)为代表的细胞还原氧化状态调节CLOCK/BMAL1和NPAS2/BMAL1的转录活性,NAD还原形式能够提高NPAS2的DNA结合活性,而氧化形式降低了NPAS2的DNA结合活性[30].TCCD诱导的多聚腺苷二磷酸核糖聚合酶(TCDD inducible poly-ADP-ribose polymerase,TIPARP)是一种单ADP-核糖基转移酶,是PARP家族成员,受芳烃受体(aryl hydrocarbon receptor,AHR)调控,其详细功能及其在调节转录中的作用尚不清楚.有研究表明,TIPARP是HIF-1的靶标,充当缺氧信号转导的负反馈调节剂,能够以ADP-核糖基化依赖性方式形成球形核体并募集HIF-1α和E3泛素连接酶HUWE1,导致HIF-1α的泛素化和降解[31].此外研究[32]发现,OA中的TIPARP表达显著降低. ...
1
... 除了NAMPT,关键基因NPAS2和TIPARP在OA中发挥的作用也同样重要.神经元PAS结构域蛋白2(neuronal PAS domain protein 2,NPAS2)是一种调节哺乳动物昼夜节律的转录因子,能够通过与脑和肌肉ARNT样蛋白1(brain and muscle arnt-like protein 1,BMAL1)形成二聚体参与昼夜节律的调节[29].研究表明,以NAD(H)和NADP(H)为代表的细胞还原氧化状态调节CLOCK/BMAL1和NPAS2/BMAL1的转录活性,NAD还原形式能够提高NPAS2的DNA结合活性,而氧化形式降低了NPAS2的DNA结合活性[30].TCCD诱导的多聚腺苷二磷酸核糖聚合酶(TCDD inducible poly-ADP-ribose polymerase,TIPARP)是一种单ADP-核糖基转移酶,是PARP家族成员,受芳烃受体(aryl hydrocarbon receptor,AHR)调控,其详细功能及其在调节转录中的作用尚不清楚.有研究表明,TIPARP是HIF-1的靶标,充当缺氧信号转导的负反馈调节剂,能够以ADP-核糖基化依赖性方式形成球形核体并募集HIF-1α和E3泛素连接酶HUWE1,导致HIF-1α的泛素化和降解[31].此外研究[32]发现,OA中的TIPARP表达显著降低. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}