Congenital tooth agenesis-related EDAR variants and pedigree analysis of HED patients with two variants
LAN Rong,, DAI Qinggang, YU Kang, BIAN Xiaoling, YE Lijuan, WU Yiqun, WANG Feng,
Department of 2nd Dental Center, Shanghai Ninth People's Hospital, Shanghai Jiao Tong University School of Medicine; College of Stomatology, Shanghai Jiao Tong University; National Center for Stomatology; National Clinical Research Center for Oral Diseases; Shanghai Key Laboratory of Stomatology; Shanghai Research Institute of Stomatology, Shanghai 201999, China
Objective ·To explore EDAR (ectodysplasin A receptor) gene variants that lead to congenital tooth agenesis, and preliminarily analyze the reasons why variants in EDAR can cause both syndromic and non-syndromic tooth agenesis. Methods ·Patients with congenital tooth agenesis admitted to the Department of 2nd Dental Center, Shanghai Ninth People's Hospital, Shanghai Jiao Tong University School of Medicine and their family members were included, and genomic DNA from their peripheral blood was extracted for whole exome sequencing (WES). After preliminary screening, PolyPhen-2, Mutation Taster, and Provean were used to predict the harmfulness of potential variants. The screened variants in patients and their families were verified by Sanger sequencing. Conservation analysis of variants was performed, and Swiss-Model was used to analyze the changes in the three-dimensional structure of EDAR. The teeth and syndromic phenotype of the patients and their family members were investigated. Results ·Among the included congenital tooth agenesis patients, five patients with EDAR mutations were found, one with EDAR frameshift mutation c.368_369insC(p.L123fs) and the other four with EDAR missense mutations. Two of these four patients were diagnosed as non-syndromic tooth agenesis (NSTA), resulted from c.77C>A(p.A26E) homozygous mutation and c.380C>T(p.P127L) heterozygous mutation, respectively. The other two patients with two variants were diagnosed as hypohidrotic ectodermal dysplasia (HED). One compound heterozygous missense mutation patient carried EDAR c.77C>T(p.A26V) from her father and EDAR c.1281G>C (p.L427F) from her mother; the other patient with both EDAR and EDA mutations carried EDAR c.1138A>C(p.S380R) heterozygous mutation and EDA c.1013C>T(p.T338M) hemizygous mutation. Both variants were from his mother and were reported to be related with NSTA. Two of these missense mutations, EDAR c.1281G>C(p.L427F) and EDAR c.77C>A (p.A26E), had not been reported before. The missense mutations affected the protein's spatial conformation by altering the polarity, charge, or volume of the amino acid residues. The frameshift mutation caused a non-triplet base addition, which probably led to protein truncation or degradation. Conclusion ·Two new EDAR missense mutations are discovered. An NSTA patients with EDAR homozygous mutations and an HED patient with both EDA and EDAR mutations are reported. It expands the understanding of pathogenic mechanisms of EDAR mutations causing HED and NSTA.
Keywords:ectodermal dysplasia
;
familial tooth agenesis
;
ectodysplasin A receptor
;
ectodysplasin A
;
whole exome sequencing
LAN Rong, DAI Qinggang, YU Kang, BIAN Xiaoling, YE Lijuan, WU Yiqun, WANG Feng. Congenital tooth agenesis-related EDAR variants and pedigree analysis of HED patients with two variants. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2024, 44(6): 694-701 doi:10.3969/j.issn.1674-8115.2024.06.004
先天缺牙是由于牙胚发育障碍,造成牙齿数目减少的常见颅颌面先天发育缺陷。它在不同人群中的发病率可达1.6%~6.9%(除第三磨牙外),其发病与遗传、环境、进化等多种因素有关,遗传因素在家族性先天缺牙中起主导作用[1]。在发育过程中,牙齿的形态和数量由外异蛋白A(ectodysplasin A,EDA)、无翅型MMTV整合位点家族(wingless-type MMTV integration site family,WNT)、音猬因子(sonic hedgehog,SHH)共同调控[2]。除了影响牙齿外,这些通路也与皮肤、指甲等其他外胚层器官的发育有关。因此,以上通路的异常还可以导致少汗型外胚层发育不良(hypohidrotic ectodermal dysplasia,HED),HED患者具有毛发稀少、少汗、先天缺牙、特殊面容(鞍状鼻、眼周色素沉着、前额突出)等症状[2-3]。
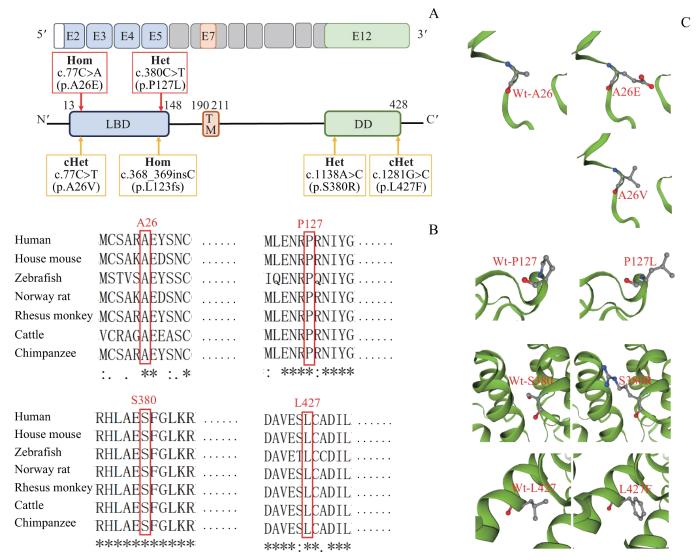
外异蛋白A受体(ectodysplasin A receptor,EDAR)作为EDA的受体,发挥着关键的信号传递作用。EDAR基因位于2号染色体q11~q13,包含12个外显子。此前有研究[4]发现,除EDA突变致病外,有1/4的HED病例都是由EDAR突变所致。EDAR基因的第2、3、4外显子编码与EDA结合的胞外配体结合域(ligand binding domain,LBD);第12外显子编码胞内死亡结构域(death domain,DD),该区域募集EDARADD进而激活下游的核因子κB(nuclear factor κB,NF-κB),介导外胚层来源组织器官的生长发育[3]。这2个区域也是突变最高发的区域,突变可能导致EDAR蛋白构象发生改变,影响蛋白稳定性,使其与EDA或EDARADD的亲和力下降[5]。
根据美国医学遗传学和基因组学学院(American College of Medical Genetics and Genomics,ACMG)的指南,突变被分类为致病性(pathogenic)、可能致病性(likely pathogenic)、不确定意义(uncertain significance)、可能良性(likely benign)、良性(benign)。
1.5 突变位点保守性分析及蛋白结构预测
通过NCBI(National Center of Biotechnology Information)数据库(https://www.ncbi.nlm.nih.gov/)获得不同物种的EDAR蛋白序列,包括人(>NP_071731.1)、小鼠(>NP_034230.1)、斑马鱼(>NP_001108536.2)、牛(>XP_005212787.1)、褐家鼠(>NP_001178828.1)、恒河猴(>XP_014968589.2)、黑猩猩(>XP_024210387.1)这7种脊椎动物的EDAR氨基酸序列,使用T-Coffee软件(https://www.ebi.ac.uk/Tools/msa/tcoffee/)进行了跨物种的保守性分析。
Note: A. Schematic diagram of wild-type EDAR protein and the localization of identified EDAR variants (red squares indicate variants found in NSTA patients, and yellow ones represent those in HED patients). B. Conservation analysis of affected amino acids among seven vertebrate species. C. Three-dimensional structure of wild-type EDAR and five missense variants (p.A26E, p.A26V, p.P127L, p.S380R, and p.L427F). E—exon; TM—transmembrane domain; Wt—wild-type.
Fig 1
Conservation analysis of EDAR variants and the structure prediction of mutant proteins
在Swiss-Model在线工具中选择序列匹配度最高的Q9R187.1.A模板进行EDAR蛋白的三维结构重建,野生型蛋白序列匹配度为91.52%,突变型匹配度为91.29%,GMQE(global model quality estimate)值为0.67。通过蛋白结构三维重建(图1C)可以看到,c.77C>T(p.A26V)、c.77C>A(p.A26E)分别将第26位氨基酸由非极性不带电荷的丙氨酸变为了侧链体积更大的缬氨酸和极性带负电的谷氨酸;c.380C>T(p.P127L)突变使得具有环状侧链结构的脯氨酸变成了支链氨基酸亮氨酸,影响其卷曲构象;c.1138A>C(p.S380R)导致侧链体积较小的丝氨酸突变为了体积较大并且带正电的精氨酸,可能对此处的α-螺旋结构造成影响;c.1281G>C(p.L427F)位点附近也存在α-螺旋结构,该突变使得亮氨酸变成了侧链体积较大且带有苯环的苯丙氨酸。
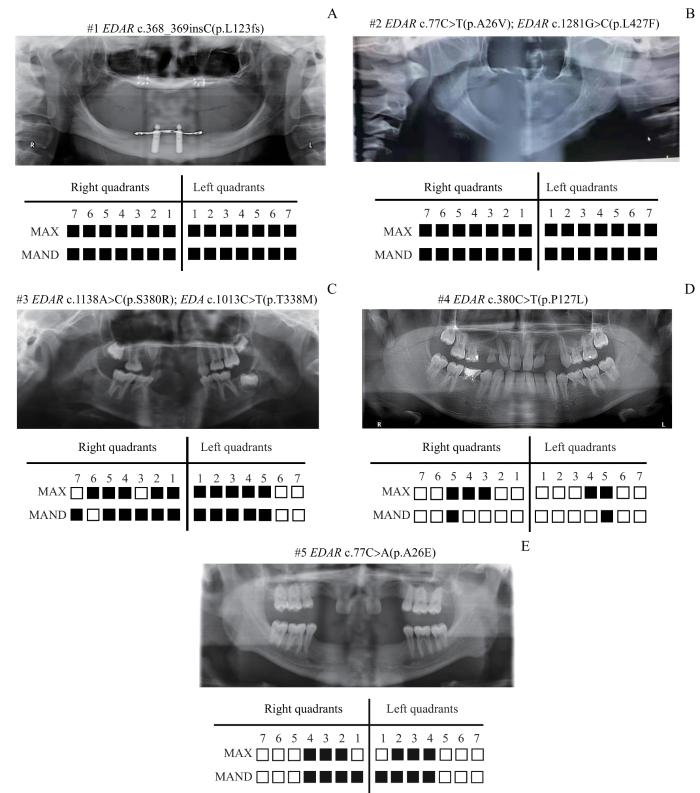
Note:A. #1 HED patient with all the teeth and/or germs missing. B. #2 HED patient with all the teeth and/or germs missing. C. #3 HED patient with 21 teeth and/or germs missing, and 6 deciduous teeth retained. D. #4 NSTA patient with 7 teeth and/or germs missing, and 5 deciduous teeth retained. E. #5 NSTA patient with 14 teeth missing.
Fig 2
Permanent teeth and/or permanent teeth germs missing in congenital tooth agenesis patients with EDAR variants
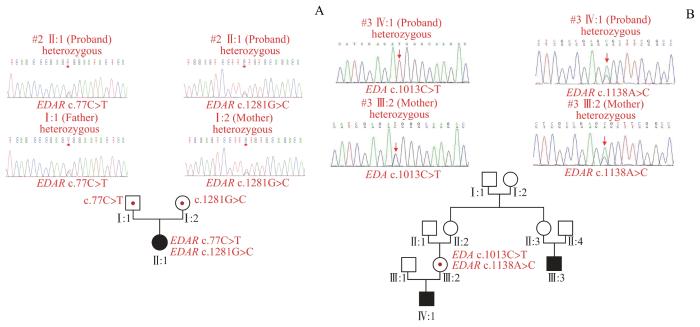
Note: A. #2 proband carried compound heterozygous missense mutation, EDAR c.77C>T(p.A26V) from her father and EDAR c.1281G>C(p.L427F) from her mother; B. #3 proband carried EDAR c.1138A>C(p.S380R) heterozygous mutation and EDA c.1013C>T(p.T338M) hemizygous mutation, both from his mother. Squares indicate males, and circles indicate females. Filled squares/circles represent individuals with tooth agenesis, and empty ones represent unaffected subjects. Red knots mean that variants were verified by Sanger sequencing.
Fig 3
Pedigree analysis and Sanger sequencing of two HED patients with two variants
#3先证者携带来自其母亲的EDAR c.1138A>C(p.S380R)杂合突变和EDA c.1013C>T(p.T338M)半合子突变,其临床表现为多牙缺失、少汗、少毛,但其母亲并无症状。以往报道认为EDAR c.1138A>C(p.S380R)杂合突变可能与NSTA相关[10];EDA c.1013C>T(p.T338M)也与NSTA有关,并且因其位于X染色体,患者通常为男性[11]。这种2个突变不单独致病但发生在同一个体就会引发疾病的现象,在遗传学中被称为“合成致死”(synthetic lethality),用于解释2个不同基因的突变具有协同致病效应[22-23]。这是首次在EDA通路中发现2个基因的突变会加重患者临床症状。此前研究[24]报道了WNT通路中Wnt蛋白10A(Wnt family member 10A,WNT10A)和低密度脂蛋白受体6(low-density lipoprotein receptor-related protein 6,LRP6)基因突变也会协同产生更严重的缺牙表型。这也说明,NSTA和HED很可能是严重程度不同的同种疾病。
The study was designed and supervised by WANG Feng and WU Yiqun. The manuscript was drafted and revised by LAN Rong and WANG Feng. The data were collated by LAN Rong, DAI Qinggang, YU Kang and BIAN Xiaoling. The clinical cases were collected by DAI Qinggang and YE Lijuan. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors have no conflict of interest relevant to this study.
AL-ANI A H, ANTOUN J S, THOMSON W M, et al. Hypodontia: an update on its etiology, classification, and clinical management[J]. Biomed Res Int, 2017, 2017: 9378325.
CHASSAING N, BOURTHOUMIEU S, COSSEE M, et al. Mutations in EDAR account for one-quarter of non-ED1-related hypohidrotic ectodermal dysplasia[J]. Hum Mutat, 2006, 27(3): 255-259.
GAO Y Z, JIANG X H, WEI Z, et al. The EDA/EDAR/NF-κB pathway in non-syndromic tooth agenesis: a genetic perspective[J]. Front Genet, 2023, 14: 1168538.
PARVEEN A, KHAN S A, MIRZA M U, et al. Deleterious variants in WNT10A, EDAR, and EDA causing isolated and syndromic tooth agenesis: a structural perspective from molecular dynamics simulations[J]. Int J Mol Sci, 2019, 20(21): 5282.
PLAISANCIÉ J, BAILLEUL-FORESTIER I, GASTON V, et al. Mutations in WNT10A are frequently involved in oligodontia associated with minor signs of ectodermal dysplasia[J]. Am J Med Genet A, 2013, 161A(4): 671-678.
ZHANG L T, YU M, WONG S W, et al. Comparative analysis of rare EDAR mutations and tooth agenesis pattern in EDAR- and EDA-associated nonsyndromic oligodontia[J]. Hum Mutat, 2020, 41(11): 1957-1966.
LI S F, LI J H, CHENG J, et al. Non-syndromic tooth agenesis in two Chinese families associated with novel missense mutations in the TNF domain of EDA (ectodysplasin A)[J]. PLoS One, 2008, 3(6): e2396.
ZHANG L T. Functional study and susceptible tooth position of non-syndromic tooth agenesis caused by EDAR mutations[D]. Beijing: Peking University, 2021.
ZENG B H, ZHAO Q, LI S J, et al. Novel EDA or EDAR mutations identified in patients with X-linked hypohidrotic ectodermal dysplasia or non-syndromic tooth agenesis[J]. Genes, 2017, 8(10): 259.
FOURNIER B P, BRUNEAU M H, TOUPENAY S, et al. Patterns of dental agenesis highlight the nature of the causative mutated genes[J]. J Dent Res, 2018, 97(12): 1306-1316.
CHEN Y T, LIU H C, HAN D, et al. Association between EDAR polymorphisms and non-syndromic tooth agenesis in the Chinese Han population[J]. Chin J Dent Res, 2017, 20(3): 153-159.
ZHANG H Y, YANG L Y, MENG L Q, et al. A novel EDAR mutation identified in nonsyndromic oligodontia patients by whole exome sequencing[J]. Journal of Practical Stomatology, 2021, 7(6): 783-789.
LIU X Y, ZHU J X, ZHAO Y M. Whole exome sequencing and analysis of hypohidrotic ectodermal dysplasia patients[J]. Chinese Journal of Stomatology, 2022, 57(2): 154-160.
CASAL M L, LEWIS J R, MAULDIN E A, et al. Significant correction of disease after postnatal administration of recombinant ectodysplasin A in canine X-linked ectodermal dysplasia[J]. Am J Hum Genet, 2007, 81(5): 1050-1056.
YU K, HUANG C H, WAN F T, et al. Structural insights into pathogenic mechanism of hypohidrotic ectodermal dysplasia caused by ectodysplasin A variants[J]. Nat Commun, 2023, 14(1): 767.
VAN DER HOUT A H, OUDESLUIJS G G, VENEMA A, et al. Mutation screening of the ectodysplasin-A receptor gene EDAR in hypohidrotic ectodermal dysplasia[J]. Eur J Hum Genet, 2008, 16(6): 673-679.
HAN Y, WANG X L, ZHENG L Y, et al. Pathogenic EDA mutations in Chinese Han families with hypohidrotic ectodermal dysplasia and genotype-phenotype: a correlation analysis[J]. Front Genet, 2020, 11: 21.
... #3先证者携带来自其母亲的EDAR c.1138A>C(p.S380R)杂合突变和EDA c.1013C>T(p.T338M)半合子突变,其临床表现为多牙缺失、少汗、少毛,但其母亲并无症状.以往报道认为EDAR c.1138A>C(p.S380R)杂合突变可能与NSTA相关[10];EDA c.1013C>T(p.T338M)也与NSTA有关,并且因其位于X染色体,患者通常为男性[11].这种2个突变不单独致病但发生在同一个体就会引发疾病的现象,在遗传学中被称为“合成致死”(synthetic lethality),用于解释2个不同基因的突变具有协同致病效应[22-23].这是首次在EDA通路中发现2个基因的突变会加重患者临床症状.此前研究[24]报道了WNT通路中Wnt蛋白10A(Wnt family member 10A,WNT10A)和低密度脂蛋白受体6(low-density lipoprotein receptor-related protein 6,LRP6)基因突变也会协同产生更严重的缺牙表型.这也说明,NSTA和HED很可能是严重程度不同的同种疾病. ...
... #3先证者携带来自其母亲的EDAR c.1138A>C(p.S380R)杂合突变和EDA c.1013C>T(p.T338M)半合子突变,其临床表现为多牙缺失、少汗、少毛,但其母亲并无症状.以往报道认为EDAR c.1138A>C(p.S380R)杂合突变可能与NSTA相关[10];EDA c.1013C>T(p.T338M)也与NSTA有关,并且因其位于X染色体,患者通常为男性[11].这种2个突变不单独致病但发生在同一个体就会引发疾病的现象,在遗传学中被称为“合成致死”(synthetic lethality),用于解释2个不同基因的突变具有协同致病效应[22-23].这是首次在EDA通路中发现2个基因的突变会加重患者临床症状.此前研究[24]报道了WNT通路中Wnt蛋白10A(Wnt family member 10A,WNT10A)和低密度脂蛋白受体6(low-density lipoprotein receptor-related protein 6,LRP6)基因突变也会协同产生更严重的缺牙表型.这也说明,NSTA和HED很可能是严重程度不同的同种疾病. ...
... #3先证者携带来自其母亲的EDAR c.1138A>C(p.S380R)杂合突变和EDA c.1013C>T(p.T338M)半合子突变,其临床表现为多牙缺失、少汗、少毛,但其母亲并无症状.以往报道认为EDAR c.1138A>C(p.S380R)杂合突变可能与NSTA相关[10];EDA c.1013C>T(p.T338M)也与NSTA有关,并且因其位于X染色体,患者通常为男性[11].这种2个突变不单独致病但发生在同一个体就会引发疾病的现象,在遗传学中被称为“合成致死”(synthetic lethality),用于解释2个不同基因的突变具有协同致病效应[22-23].这是首次在EDA通路中发现2个基因的突变会加重患者临床症状.此前研究[24]报道了WNT通路中Wnt蛋白10A(Wnt family member 10A,WNT10A)和低密度脂蛋白受体6(low-density lipoprotein receptor-related protein 6,LRP6)基因突变也会协同产生更严重的缺牙表型.这也说明,NSTA和HED很可能是严重程度不同的同种疾病. ...
1
... #3先证者携带来自其母亲的EDAR c.1138A>C(p.S380R)杂合突变和EDA c.1013C>T(p.T338M)半合子突变,其临床表现为多牙缺失、少汗、少毛,但其母亲并无症状.以往报道认为EDAR c.1138A>C(p.S380R)杂合突变可能与NSTA相关[10];EDA c.1013C>T(p.T338M)也与NSTA有关,并且因其位于X染色体,患者通常为男性[11].这种2个突变不单独致病但发生在同一个体就会引发疾病的现象,在遗传学中被称为“合成致死”(synthetic lethality),用于解释2个不同基因的突变具有协同致病效应[22-23].这是首次在EDA通路中发现2个基因的突变会加重患者临床症状.此前研究[24]报道了WNT通路中Wnt蛋白10A(Wnt family member 10A,WNT10A)和低密度脂蛋白受体6(low-density lipoprotein receptor-related protein 6,LRP6)基因突变也会协同产生更严重的缺牙表型.这也说明,NSTA和HED很可能是严重程度不同的同种疾病. ...
1
... #3先证者携带来自其母亲的EDAR c.1138A>C(p.S380R)杂合突变和EDA c.1013C>T(p.T338M)半合子突变,其临床表现为多牙缺失、少汗、少毛,但其母亲并无症状.以往报道认为EDAR c.1138A>C(p.S380R)杂合突变可能与NSTA相关[10];EDA c.1013C>T(p.T338M)也与NSTA有关,并且因其位于X染色体,患者通常为男性[11].这种2个突变不单独致病但发生在同一个体就会引发疾病的现象,在遗传学中被称为“合成致死”(synthetic lethality),用于解释2个不同基因的突变具有协同致病效应[22-23].这是首次在EDA通路中发现2个基因的突变会加重患者临床症状.此前研究[24]报道了WNT通路中Wnt蛋白10A(Wnt family member 10A,WNT10A)和低密度脂蛋白受体6(low-density lipoprotein receptor-related protein 6,LRP6)基因突变也会协同产生更严重的缺牙表型.这也说明,NSTA和HED很可能是严重程度不同的同种疾病. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}