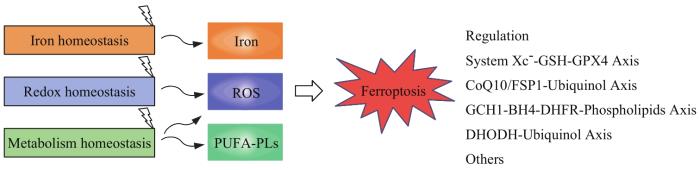
Ferroptosis is a form of cell death that results from accumulation of lipid reactive oxygen species, marked by iron-dependent oxidative damage of phospholipids. Accumulation of iron and lipid hydroperoxides are hallmarks of ferroptosis. Diverse biological contexts, including iron handling, redox homeostasis, imbalance of lipid synthesis, participate in ferroptosis. Mechanistically, pathways of ferroptosis regulation involving System Xc--glutathione (GSH)-glutathione peroxidase 4 (GPX4) axis, coenzyme Q10 (CoQ10)-ferroptosis suppressor protein 1 (FSP1)-ubiquinol axis, GTP cyclohydrolase 1 (GCH1)-tetrahydrobiopterin (BH4)-dihydrofolate reductase (DHFR) axis, dehydrogenase (DHODH)-ubiquinol axis have been discovered. Ferroptosis has been implicated in multiple liver diseases, such as hepatocellular carcinoma, liver ischemia-reperfusion injury, steatohepatitis, liver fibrosis, cirrhosis and liver metabolic diseases. Elucidating its mechanism offers various tractable nodes for therapeutic intervention. Here, we summarize insights into the molecular characteristics, biological processes, regulatory pathways of ferroptosis and its recent advances in the treatment of liver diseases.
CHEN Chen, CHENG Zhuoan, WANG Cun, XIA Qiang. Research progress in ferroptosis regulation in the treatment of liver diseases. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(3): 365-373 doi:10.3969/j.issn.1674-8115.2023.03.013
过量积聚的脂质发生过氧化是铁死亡的重要标志之一,脂肪酸代谢失衡是影响铁死亡的一个重要因素。铁死亡过程中过氧化的底物是具有多不饱和羟基尾的磷脂。由于含有双烯氢原子,PUFA容易发生脂质过氧化,即在铁依赖性酶和不稳定铁作用下使用氧分子发生过氧化反应,产生脂质过氧化物(PL-PUFA-OOH)[19]。PL-PUFA-OOH的积累最终会破坏细胞器和细胞膜结构,因此PUFA-PLs含量越高的膜越容易发生过氧化。花生四烯酸(C20∶4)或其延伸产物肾上腺酸(C22∶4)的磷脂酰乙醇胺(phosphatidyl ethanolamine,PEs)是氧化驱动铁死亡的关键磷脂[20]。研究者[21]筛选发现了两种与脂代谢相关的膜重塑酶,即乙酰辅酶A合成酶长链家族成员4(acyl-CoA synthetase long chain family member 4,ACSL4)和磷脂酰胆碱酰基转移酶3(lysophosphatidylcholine acyltransferase 3,LPCAT3)对PUFA-PLs的产生发挥重要作用。ACSL4可以连接长链PUFAs和辅酶A,其产物可被重新酯化为磷脂,增加了脂质和膜中的长链PUFAs掺入量[22]。在多种肿瘤中ACSL4的表达量都与细胞对铁死亡诱导剂的敏感性相关。LPCAT3是LPLAT异构体的主要存在形式,对PUFA具有特异性,可调节磷脂和缩醛磷脂中PUFA的含量[23]。
抑制脂质过氧化系统可抑制铁死亡:如谷胱甘肽过氧化物酶4(glutathione peroxidase 4,GPX4)可参与阻断磷脂过氧化到相应的磷脂醇从而抑制铁死亡,自由基捕获抗氧化剂(radical-trapping antioxidants,RTA)和铁死亡抑制蛋白1(ferroptosis suppressor protein 1,FSP1)等参与终止自由基驱动的脂质过氧化发展阶段从而抑制铁死亡[24-25]。脂氧酶(lipoxygenases,LOXs)可以直接氧化膜中的PUFAs和含PUFA的脂质[26],但其在铁死亡中的具体作用还不清楚。GTP环化水解酶1(GTP cyclohydrolase 1,GCH1)调控的四氢生物蝶呤(tetrahydrobiopterin,BH4)合成可以选择性抑制多不饱和脂肪酸尾部消耗磷脂,造成脂质重塑从而抑制铁死亡[27]。
System Xc-是由xCT[溶质运载蛋白7家族成员11(solute carrier family 7 member 11,SLC7A11)]和4F2重链(SLC3A2)两个亚基组成的异源二聚体,可向细胞内转入胱氨酸,同时释放1个谷氨酸到胞外。SLC7A11的表达和活性受到多种因素调节,包括肿瘤蛋白P53(tumor protein P53,TP53)、核转录因子E2相关因子2(nuclear factor erythroid 2-like-2,NFE2L2)/NRF2、乳腺癌易感基因1(breast cancer susceptibility gene 1,BRCA1)相关蛋白1、黏蛋白1(mucin1,MUC1)、Beclin1(BECN1)等,进而影响GSH的水平。抑制SLC7A11通路是诱导铁死亡的重要机制之一。GSH是由谷氨酸(glutamic acid,Glu)、半胱氨酸(cysteine,Cys)、甘氨酸(glycine,Gly)组成的三肽,其中的半胱氨酸巯基是其作为抗氧化剂的主要功能基团。组成GSH的半胱氨酸,可通过转硫途径从甲硫氨酸中获得,也可通过System Xc-从细胞外的胱氨酸中获得。GPX4是一种硒蛋白,是抗氧化的关键酶,可以直接把磷脂过氧化物还原为磷脂过氧化氢[31-32]。一旦膜发生氧化损伤,细胞会启动至少2种机制进行修复:第一种是激活特异性的酶[如GPX4、凋亡诱导因子线粒体相关2(apoptosis inducing factor mitochondrial 2,AIFM2)等]来限制脂质过氧化,第二种是使用囊泡转运、胞吞胞吐修复损伤的膜。硒元素可通过转录因子AP-2 gamma和特异性蛋白1上调GPX4的表达[33]。目前,人们公认System Xc--GSH-GPX4轴是铁死亡过程中主要的负向调控轴。
他汀类药物(氟伐他汀、洛伐他汀酸、辛伐他汀等)通过减少甲羟戊酸途径中异戊烯焦磷酸(isopentenyl pyrophosphate,IPP)产生,减少硒蛋白和CoQ10生物合成,从而促进间质型细胞发生铁死亡[48]。Brequinar是一种喹啉羧酸衍生物,最初开发用作抗癌药物,它可抑制DHODH诱导铁死亡[36]。FSP1为铁死亡抑制基因,FSP1 inhibitor(iFSP1)抑制FSP1,与RSL3协同诱导铁死亡[25]。甲氨蝶呤抑制DHFR,协同抑制GPX4从而诱导铁死亡[35]。青蒿素类如蒿甲醚、青蒿琥酯、双氢青蒿素会影响许多通路进而影响铁死亡。2015年有研究[49]报道了青蒿琥酯在Kirsten鼠类肉瘤(Kirsten rat sarcoma,KRAS)转换的胰腺导管腺癌细胞中诱导铁死亡;2019年有报道[9]发现,双氢青蒿素通过促进铁蛋白吞噬、增加游离铁促进铁死亡,且证实在治疗急性髓系白血病中可发挥作用。纳米材料也为铁死亡诱导剂的研究提供了新的可能。
肝细胞癌(hepatocellular carcinoma,HCC)是一种多基因参与、多因素介导、病理机制复杂的恶性肿瘤,铁死亡治疗肝细胞癌是一个新的研究方向。生物信息学分析[51]发现使用铁死亡相关基因构建的模型可有效区分肝细胞癌患者和健康对照,且高风险组预后更差。高铁饮食会增加HCC患病风险。Erastin诱导铁死亡可抑制HCC增殖和进展[51]。HCC治疗药物索拉非尼可抑制SLC7A11促进肝癌细胞铁死亡。IFN-γ通过酪氨酸蛋白激酶/信号转导子和转录激活子(Janus kinase/signal transducer and activator of transcription,JAK/STAT)信号通路抑制System Xc-,使HCC对铁死亡敏感[52]。
NPD由于遗传性的神经鞘磷脂酶缺乏造成溶酶体功能异常,导致鞘磷脂在包括肝脏在内的多个器官系统中积累。NPD C型(NPD type C,NPC)患者的成纤维细胞中ROS和脂质过氧化水平较正常对照明显升高[65],患者血清中氧化应激相关标志物上升[66],CoQ10减少[67]。抑制铁死亡相关损伤也许会为NPD的治疗提供新思路。
The manuscript was drafted by CHEN Chen and CHENG Zhuoan, and revised by WANG Cun and XIA Qiang. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
All authors disclose no relevant conflict of interests.
DOLMA S, LESSNICK S L, HAHN W C, et al. Identification of genotype-selective antitumor agents using synthetic lethal chemical screening in engineered human tumor cells[J]. Cancer Cell, 2003, 3(3): 285-296.
YANG W S, STOCKWELL B R. Synthetic lethal screening identifies compounds activating iron-dependent, nonapoptotic cell death in oncogenic-RAS-harboring cancer cells[J]. Chem Biol, 2008, 15(3): 234-245.
NEMETH E, TUTTLE M S, POWELSON J, et al. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization[J]. Science, 2004, 306(5704): 2090-2093.
ALVAREZ S W, SVIDERSKIY V O, TERZI E M, et al. NFS1 undergoes positive selection in lung tumours and protects cells from ferroptosis[J]. Nature, 2017, 551(7682): 639-643.
TERZI E M, SVIDERSKIY V O, ALVAREZ S W, et al. Iron-sulfur cluster deficiency can be sensed by IRP2 and regulates iron homeostasis and sensitivity to ferroptosis independent of IRP1 and FBXL5[J]. Sci Adv, 2021, 7(22): eabg4302.
DU J, WANG T, LI Y, et al. DHA inhibits proliferation and induces ferroptosis of leukemia cells through autophagy dependent degradation of ferritin[J]. Free Radic Biol Med, 2019, 131: 356-369.
MA S, HENSON E S, CHEN Y, et al. Ferroptosis is induced following siramesine and lapatinib treatment of breast cancer cells[J]. Cell Death Dis, 2016, 7(7): e2307.
YANG W H, HUANG Z Q, WU J L, et al. A TAZ-ANGPTL4-NOX2 axis regulates ferroptotic cell death and chemoresistance in epithelial ovarian cancer[J]. Mol Cancer Res, 2020, 18(1): 79-90.
CHEN X, XU S, ZHAO C, et al. Role of TLR4/NADPH oxidase 4 pathway in promoting cell death through autophagy and ferroptosis during heart failure[J]. Biochem Biophys Res Commun, 2019, 516(1): 37-43.
YANG W H, DING C C, SUN T, et al. The hippo pathway effector TAZ regulates ferroptosis in renal cell carcinoma[J]. Cell Rep, 2019, 28(10): 2501-2508.e4.
XIE Y, ZHU S, SONG X, et al. The tumor suppressor p53 limits ferroptosis by blocking DPP4 activity[J]. Cell Rep, 2017, 20(7): 1692-1704.
ZOU Y L, LI H X, GRAHAM E T, et al. Cytochrome P450 oxidoreductase contributes to phospholipid peroxidation in ferroptosis[J]. Nat Chem Biol, 2020, 16(3): 302-309.
DIXON S J, WINTER G E, MUSAVI L S, et al. Human haploid cell genetics reveals roles for lipid metabolism genes in nonapoptotic cell death[J]. ACS Chem Biol, 2015, 10(7): 1604-1609.
LAGROST L, MASSON D. The expanding role of lyso-phosphatidylcholine acyltransferase-3 (LPCAT3), a phospholipid remodeling enzyme, in health and disease[J]. Curr Opin Lipidol, 2022, 33(3): 193-198.
BERSUKER K, HENDRICKS J M, LI Z P, et al. The CoQ oxidoreductase FSP1 acts parallel to GPX4 to inhibit ferroptosis[J]. Nature, 2019, 575(7784): 688-692.
KRAFT V A N, BEZJIAN C T, PFEIFFER S, et al. GTP cyclohydrolase 1/tetrahydrobiopterin counteract ferroptosis through lipid remodeling[J]. ACS Cent Sci, 2020, 6(1): 41-53.
SHIN D, LEE J, YOU J H, et al. Dihydrolipoamide dehydrogenase regulates cystine deprivation-induced ferroptosis in head and neck cancer[J]. Redox Biol, 2020, 30: 101418.
URSINI F, MAIORINO M, VALENTE M, et al. Purification from pig liver of a protein which protects liposomes and biomembranes from peroxidative degradation and exhibits glutathione peroxidase activity on phosphatidylcholine hydroperoxides[J]. Biochim Biophys Acta, 1982, 710(2): 197-211.
ALIM I, CAULFIELD JT, CHEN Y, et al. Selenium drives a transcriptional adaptive program to block ferroptosis and treat stroke[J]. Cell, 2019, 177(5): 1262-1279.e25.
VENKATESH D, O'BRIEN N A, ZANDKARIMI F, et al. MDM2 and MDMX promote ferroptosis by PPARα-mediated lipid remodeling[J]. Genes Dev, 2020, 34(7/8): 526-543.
SOULA M, WEBER R A, ZILKA O, et al. Metabolic determinants of cancer cell sensitivity to canonical ferroptosis inducers[J]. Nat Chem Biol, 2020, 16(12): 1351-1360.
CHEN D, FAN Z, RAUH M, et al. ATF4 promotes angiogenesis and neuronal cell death and confers ferroptosis in a xCT-dependent manner[J]. Oncogene, 2017, 36(40): 5593-5608.
WU J, MINIKES A M, GAO M H, et al. Intercellular interaction dictates cancer cell ferroptosis via NF2-YAP signalling[J]. Nature, 2019, 572(7769): 402-406.
GAO R Z, KALATHUR R K R, COTO-LLERENA M, et al. YAP/TAZ and ATF4 drive resistance to Sorafenib in hepatocellular carcinoma by preventing ferroptosis[J]. EMBO Mol Med, 2021, 13(12): e14351.
SINGHAL R, MITTA S R, DAS N K, et al. HIF-2α activation potentiates oxidative cell death in colorectal cancers by increasing cellular iron[J]. J Clin Invest, 2021, 131(12): e143691.
HANGAUER M J, VISWANATHAN V S, RYAN M J, et al. Drug-tolerant persister cancer cells are vulnerable to GPX4 inhibition[J]. Nature, 2017, 551(7679): 247-250.
GASCHLER M M, ANDIA A A, LIU H R, et al. FINO2 initiates ferroptosis through GPX4 inactivation and iron oxidation[J]. Nat Chem Biol, 2018, 14(5): 507-515.
LOUANDRE C, EZZOUKHRY Z, GODIN C, et al. Iron-dependent cell death of hepatocellular carcinoma cells exposed to sorafenib[J]. Int J Cancer, 2013, 133(7): 1732-1742.
VISWANATHAN V S, RYAN M J, DHRUV H D, et al. Dependency of a therapy-resistant state of cancer cells on a lipid peroxidase pathway[J]. Nature, 2017, 547(7664): 453-457.
ELING N, REUTER L, HAZIN J, et al. Identification of artesunate as a specific activator of ferroptosis in pancreatic cancer cells[J]. Oncoscience, 2015, 2(5): 517-532.
ANGELI J P F, SCHNEIDER M, PRONETH B, et al. Inactivation of the ferroptosis regulator Gpx4 triggers acute renal failure in mice[J]. Nat Cell Biol, 2014, 16(12): 1180-1191.
TANG B F, ZHU J Y, LI J, et al. The ferroptosis and iron-metabolism signature robustly predicts clinical diagnosis, prognosis and immune microenvironment for hepatocellular carcinoma[J]. Cell Commun Signal, 2020, 18(1): 174.
KONG R, WANG N, HAN W, et al. IFNγ-mediated repression of system xc- drives vulnerability to induced ferroptosis in hepatocellular carcinoma cells[J]. J Leukoc Biol, 2021, 110(2): 301-314.
WU S, YANG J, SUN G L, et al. Macrophage extracellular traps aggravate iron overload-related liver ischaemia/reperfusion injury[J]. Br J Pharmacol, 2021, 178(18): 3783-3796.
YAMADA N, KARASAWA T, WAKIYA T, et al. Iron overload as a risk factor for hepatic ischemia-reperfusion injury in liver transplantation: potential role of ferroptosis[J]. Am J Transplant, 2020, 20(6): 1606-1618.
LI Y, FENG D C, WANG Z Y, et al. Ischemia-induced ACSL4 activation contributes to ferroptosis-mediated tissue injury in intestinal ischemia/reperfusion[J]. Cell Death Differ, 2019, 26(11): 2284-2299.
LI Z Y, AGELLON L B, ALLEN T M, et al. The ratio of phosphatidylcholine to phosphatidylethanolamine influences membrane integrity and steatohepatitis[J]. Cell Metab, 2006, 3(5): 321-331.
NELSON J E, WILSON L, BRUNT E M, et al. Relationship between the pattern of hepatic iron deposition and histological severity in nonalcoholic fatty liver disease[J]. Hepatology, 2011, 53(2): 448-457.
ZHANG Z L, YAO Z, WANG L, et al. Activation of ferritinophagy is required for the RNA-binding protein ELAVL1/HuR to regulate ferroptosis in hepatic stellate cells[J]. Autophagy, 2018, 14(12): 2083-2103.
WU A M, FENG B, YU J, et al. Fibroblast growth factor 21 attenuates iron overload-induced liver injury and fibrosis by inhibiting ferroptosis[J]. Redox Biol, 2021, 46: 102131.
VÁZQUEZ M C, BALBOA E, ALVAREZ A R, et al. Oxidative stress: a pathogenic mechanism for Niemann-Pick type C disease[J]. Oxid Med Cell Longev, 2012, 2012: 205713.
... 过量积聚的脂质发生过氧化是铁死亡的重要标志之一,脂肪酸代谢失衡是影响铁死亡的一个重要因素.铁死亡过程中过氧化的底物是具有多不饱和羟基尾的磷脂.由于含有双烯氢原子,PUFA容易发生脂质过氧化,即在铁依赖性酶和不稳定铁作用下使用氧分子发生过氧化反应,产生脂质过氧化物(PL-PUFA-OOH)[19].PL-PUFA-OOH的积累最终会破坏细胞器和细胞膜结构,因此PUFA-PLs含量越高的膜越容易发生过氧化.花生四烯酸(C20∶4)或其延伸产物肾上腺酸(C22∶4)的磷脂酰乙醇胺(phosphatidyl ethanolamine,PEs)是氧化驱动铁死亡的关键磷脂[20].研究者[21]筛选发现了两种与脂代谢相关的膜重塑酶,即乙酰辅酶A合成酶长链家族成员4(acyl-CoA synthetase long chain family member 4,ACSL4)和磷脂酰胆碱酰基转移酶3(lysophosphatidylcholine acyltransferase 3,LPCAT3)对PUFA-PLs的产生发挥重要作用.ACSL4可以连接长链PUFAs和辅酶A,其产物可被重新酯化为磷脂,增加了脂质和膜中的长链PUFAs掺入量[22].在多种肿瘤中ACSL4的表达量都与细胞对铁死亡诱导剂的敏感性相关.LPCAT3是LPLAT异构体的主要存在形式,对PUFA具有特异性,可调节磷脂和缩醛磷脂中PUFA的含量[23]. ...
1
... 过量积聚的脂质发生过氧化是铁死亡的重要标志之一,脂肪酸代谢失衡是影响铁死亡的一个重要因素.铁死亡过程中过氧化的底物是具有多不饱和羟基尾的磷脂.由于含有双烯氢原子,PUFA容易发生脂质过氧化,即在铁依赖性酶和不稳定铁作用下使用氧分子发生过氧化反应,产生脂质过氧化物(PL-PUFA-OOH)[19].PL-PUFA-OOH的积累最终会破坏细胞器和细胞膜结构,因此PUFA-PLs含量越高的膜越容易发生过氧化.花生四烯酸(C20∶4)或其延伸产物肾上腺酸(C22∶4)的磷脂酰乙醇胺(phosphatidyl ethanolamine,PEs)是氧化驱动铁死亡的关键磷脂[20].研究者[21]筛选发现了两种与脂代谢相关的膜重塑酶,即乙酰辅酶A合成酶长链家族成员4(acyl-CoA synthetase long chain family member 4,ACSL4)和磷脂酰胆碱酰基转移酶3(lysophosphatidylcholine acyltransferase 3,LPCAT3)对PUFA-PLs的产生发挥重要作用.ACSL4可以连接长链PUFAs和辅酶A,其产物可被重新酯化为磷脂,增加了脂质和膜中的长链PUFAs掺入量[22].在多种肿瘤中ACSL4的表达量都与细胞对铁死亡诱导剂的敏感性相关.LPCAT3是LPLAT异构体的主要存在形式,对PUFA具有特异性,可调节磷脂和缩醛磷脂中PUFA的含量[23]. ...
1
... 过量积聚的脂质发生过氧化是铁死亡的重要标志之一,脂肪酸代谢失衡是影响铁死亡的一个重要因素.铁死亡过程中过氧化的底物是具有多不饱和羟基尾的磷脂.由于含有双烯氢原子,PUFA容易发生脂质过氧化,即在铁依赖性酶和不稳定铁作用下使用氧分子发生过氧化反应,产生脂质过氧化物(PL-PUFA-OOH)[19].PL-PUFA-OOH的积累最终会破坏细胞器和细胞膜结构,因此PUFA-PLs含量越高的膜越容易发生过氧化.花生四烯酸(C20∶4)或其延伸产物肾上腺酸(C22∶4)的磷脂酰乙醇胺(phosphatidyl ethanolamine,PEs)是氧化驱动铁死亡的关键磷脂[20].研究者[21]筛选发现了两种与脂代谢相关的膜重塑酶,即乙酰辅酶A合成酶长链家族成员4(acyl-CoA synthetase long chain family member 4,ACSL4)和磷脂酰胆碱酰基转移酶3(lysophosphatidylcholine acyltransferase 3,LPCAT3)对PUFA-PLs的产生发挥重要作用.ACSL4可以连接长链PUFAs和辅酶A,其产物可被重新酯化为磷脂,增加了脂质和膜中的长链PUFAs掺入量[22].在多种肿瘤中ACSL4的表达量都与细胞对铁死亡诱导剂的敏感性相关.LPCAT3是LPLAT异构体的主要存在形式,对PUFA具有特异性,可调节磷脂和缩醛磷脂中PUFA的含量[23]. ...
2
... 过量积聚的脂质发生过氧化是铁死亡的重要标志之一,脂肪酸代谢失衡是影响铁死亡的一个重要因素.铁死亡过程中过氧化的底物是具有多不饱和羟基尾的磷脂.由于含有双烯氢原子,PUFA容易发生脂质过氧化,即在铁依赖性酶和不稳定铁作用下使用氧分子发生过氧化反应,产生脂质过氧化物(PL-PUFA-OOH)[19].PL-PUFA-OOH的积累最终会破坏细胞器和细胞膜结构,因此PUFA-PLs含量越高的膜越容易发生过氧化.花生四烯酸(C20∶4)或其延伸产物肾上腺酸(C22∶4)的磷脂酰乙醇胺(phosphatidyl ethanolamine,PEs)是氧化驱动铁死亡的关键磷脂[20].研究者[21]筛选发现了两种与脂代谢相关的膜重塑酶,即乙酰辅酶A合成酶长链家族成员4(acyl-CoA synthetase long chain family member 4,ACSL4)和磷脂酰胆碱酰基转移酶3(lysophosphatidylcholine acyltransferase 3,LPCAT3)对PUFA-PLs的产生发挥重要作用.ACSL4可以连接长链PUFAs和辅酶A,其产物可被重新酯化为磷脂,增加了脂质和膜中的长链PUFAs掺入量[22].在多种肿瘤中ACSL4的表达量都与细胞对铁死亡诱导剂的敏感性相关.LPCAT3是LPLAT异构体的主要存在形式,对PUFA具有特异性,可调节磷脂和缩醛磷脂中PUFA的含量[23]. ...
... 过量积聚的脂质发生过氧化是铁死亡的重要标志之一,脂肪酸代谢失衡是影响铁死亡的一个重要因素.铁死亡过程中过氧化的底物是具有多不饱和羟基尾的磷脂.由于含有双烯氢原子,PUFA容易发生脂质过氧化,即在铁依赖性酶和不稳定铁作用下使用氧分子发生过氧化反应,产生脂质过氧化物(PL-PUFA-OOH)[19].PL-PUFA-OOH的积累最终会破坏细胞器和细胞膜结构,因此PUFA-PLs含量越高的膜越容易发生过氧化.花生四烯酸(C20∶4)或其延伸产物肾上腺酸(C22∶4)的磷脂酰乙醇胺(phosphatidyl ethanolamine,PEs)是氧化驱动铁死亡的关键磷脂[20].研究者[21]筛选发现了两种与脂代谢相关的膜重塑酶,即乙酰辅酶A合成酶长链家族成员4(acyl-CoA synthetase long chain family member 4,ACSL4)和磷脂酰胆碱酰基转移酶3(lysophosphatidylcholine acyltransferase 3,LPCAT3)对PUFA-PLs的产生发挥重要作用.ACSL4可以连接长链PUFAs和辅酶A,其产物可被重新酯化为磷脂,增加了脂质和膜中的长链PUFAs掺入量[22].在多种肿瘤中ACSL4的表达量都与细胞对铁死亡诱导剂的敏感性相关.LPCAT3是LPLAT异构体的主要存在形式,对PUFA具有特异性,可调节磷脂和缩醛磷脂中PUFA的含量[23]. ...
... 肝细胞癌(hepatocellular carcinoma,HCC)是一种多基因参与、多因素介导、病理机制复杂的恶性肿瘤,铁死亡治疗肝细胞癌是一个新的研究方向.生物信息学分析[51]发现使用铁死亡相关基因构建的模型可有效区分肝细胞癌患者和健康对照,且高风险组预后更差.高铁饮食会增加HCC患病风险.Erastin诱导铁死亡可抑制HCC增殖和进展[51].HCC治疗药物索拉非尼可抑制SLC7A11促进肝癌细胞铁死亡.IFN-γ通过酪氨酸蛋白激酶/信号转导子和转录激活子(Janus kinase/signal transducer and activator of transcription,JAK/STAT)信号通路抑制System Xc-,使HCC对铁死亡敏感[52]. ...
... [51].HCC治疗药物索拉非尼可抑制SLC7A11促进肝癌细胞铁死亡.IFN-γ通过酪氨酸蛋白激酶/信号转导子和转录激活子(Janus kinase/signal transducer and activator of transcription,JAK/STAT)信号通路抑制System Xc-,使HCC对铁死亡敏感[52]. ...
1
... 肝细胞癌(hepatocellular carcinoma,HCC)是一种多基因参与、多因素介导、病理机制复杂的恶性肿瘤,铁死亡治疗肝细胞癌是一个新的研究方向.生物信息学分析[51]发现使用铁死亡相关基因构建的模型可有效区分肝细胞癌患者和健康对照,且高风险组预后更差.高铁饮食会增加HCC患病风险.Erastin诱导铁死亡可抑制HCC增殖和进展[51].HCC治疗药物索拉非尼可抑制SLC7A11促进肝癌细胞铁死亡.IFN-γ通过酪氨酸蛋白激酶/信号转导子和转录激活子(Janus kinase/signal transducer and activator of transcription,JAK/STAT)信号通路抑制System Xc-,使HCC对铁死亡敏感[52]. ...



{kind=link}
{kind=link}