Research progress in the pathogenesis and prognosis of ZNF384 fusion subtype acute leukemia
LI Ying,1,2, TAN Yangxia2, YIN Hongxin2, JIANG Yanling1,2, CHEN Li1, MENG Guoyu,2
1.Department of Basic Medicine, School of Medicine, Kunming University of Science and Technology, Kunming 650500, China
2.Shanghai Institute of Hematology, State Key Laboratory of Medical Genomics, National Research Center for Translational Medicine(Shanghai), RuiJin Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai 200025, China
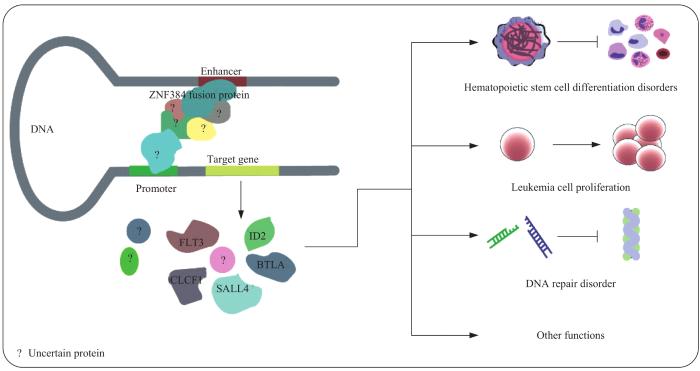
由染色体易位引起的融合基因已成为白血病的主要致病因素。锌指蛋白384(zinc finger protein 384,ZNF384)融合作为急性白血病(acute leukemia,AL)中的非典型融合亚型,在不同的年龄群体中广泛发生。ZNF384具有丰富的融合伴侣,其中E1A结合蛋白p300(E1A binding protein p300,EP300)、转录因子3(transcription factor 3,TCF3)、TATA-box binding protein associated factor 15(TAF15)的融合频率最高。这些融合蛋白均保留了完整的ZNF384结构,但融合伴侣则有不同程度的缺失,说明不同的ZNF384融合亚型之间具有相似的致AL发生发展机制。现有研究主要认为ZNF384融合蛋白通过染色质重塑调控下游蛋白的转录表达,在造血干细胞的分化、癌细胞的增殖凋亡和基因组修复中发挥潜在作用。ZNF384融合患者同时表达淋系和髓系特有的抗原,在疾病的进展中具有谱系转化特性,丰富的免疫表型给治疗方式带来了不确定性,并与融合亚型、发病年龄一起影响患者的临床结局。该文通过对近10年已发表的案例和大型队列研究进行统计归纳分析,进一步确认了ZNF384融合及其各亚型AL在现有研究背景下的发生频率,总结了已有的机制信息,并对不同治疗方式下ZNF384融合患者的预后作了简要分析,以期为后续针对这一独特亚型AL的诊疗和研究提供参考。
关键词:ZNF384融合
;
急性白血病
;
免疫表型
;
机制
;
诊断
;
预后
Abstract
Gene fusions caused by chromosomal translocations have become the main pathogenic factors that initiate leukemogenesis. Zinc finger protein 384 (ZNF384) fusion, as an atypical fusion gene in acute leukemia (AL), has widely been identified in different age groups. ZNF384 rearranged 18 genes, with E1A binding protein p300 (EP300), transcription factor 3, (TCF3), and TATA-box binding protein-associated factor 15 (TAF15) being the most common fusion partners. These fusion proteins maintain the complete structure of ZNF384, but the fusion partners are missing in varying degrees, indicating that the mechanisms behind different subtypes of carcinogenesis have similarities. The mechanism of ZNF384-rearranged AL is also being actively investigated. It is mainly believed that the fusion protein regulates the transcription and expression of downstream proteins through chromatin remodeling, and plays a potential role in the differentiation of hematopoietic stem cells, the proliferation and apoptosis of cancer cells and genome repair. Patients with ZNF384 fusions express both lymphoid and myeloid-specific antigens, which have lineage-transforming properties during disease progression. The diversity of immunophenotypes leads to ambiguity in treatment options and diverse outcomes in prognosis studies, and affects the clinical outcome of patients together with fusion subtype and age of onset. Through the statistical analysis of published cases and large-scale cohort studies in the past 10 years, the incidence of ZNF384 fusion in AL and the frequency of each fusion subtype in the context of existing research were further confirmed. The impact of different treatment methods on the prognosis of patients was analyzed, and the identified mechanisms were summarized in order to provide reference for subsequent diagnosis, treatment and research of this unique AL subtype.
LI Ying, TAN Yangxia, YIN Hongxin, JIANG Yanling, CHEN Li, MENG Guoyu. Research progress in the pathogenesis and prognosis of ZNF384 fusion subtype acute leukemia. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(5): 631-640 doi:10.3969/j.issn.1674-8115.2023.05.015
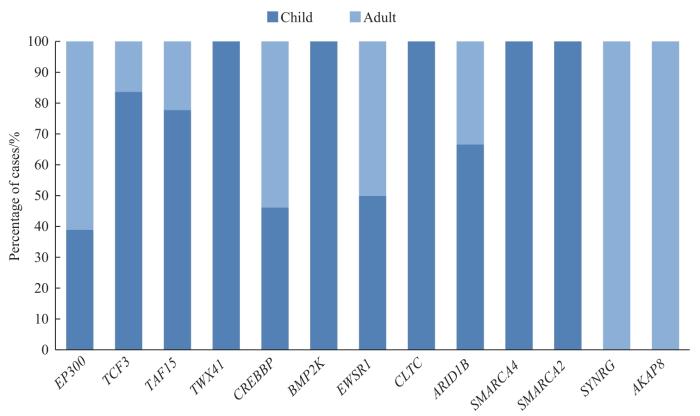
急性白血病(acute leukemia,AL)是造血干细胞恶性增殖的克隆性疾病,包括急性淋巴细胞白血病(acute lymphoblastic leukemia,ALL)、急性髓系白血病(acute myeloid leukemia,AML)、混合表型急性白血病(mixed phenotype acute leukemia,MPAL)。染色体易位已成为许多血液系统恶性肿瘤的基因组标志,异常融合基因编码的产物主要与造血分化、细胞增殖和细胞周期等机制的转录因子有关[1]。二代测序揭示了AL中存在的多种染色体易位,如肌细胞增强因子2D(myocyte enhancer factor 2D,MEF2D)融合、T细胞因子3(T cell factor 3,TCF3)-前B淋巴细胞白血病转录因子(pre-B-cell leukemia transcription factor1,PBX1)、ETS variant transcription factor 6(ETV6)-RUNX家族相关转录因子1(RUNX family transcription factor 1,RUNX1)阳性/ETV6-RUNX1样、Double homeobox 4(DUX4)融合、ZNF384融合、BCR-ABL1/Ph样和Lysine methyltransferase 2A (KMT2A)融合相关等[1-7]。其中ZNF384融合的存在较为特殊,迄今为止,已报道了18个与ZNF384发生融合的伴侣基因:E1A结合蛋白p300(E1A binding protein p300,EP300)、转录因子3(transcription factor 3,TCF3)、TATA-box binding protein associated factor 15(TAF15)、cAMP反应元件结合蛋白(cAMP-response element-binding protein,CREB)结合蛋白(CREB-binding protein,CREBBP)、EWS RNA binding protein 1(EWSR1)、转录因子AT富集区1B(AT-rich interaction domain 1B,ARID1B)、SWI/SNF related,matrix associated,actin dependent regulator of chromatin,subfamily a,member 2(SMARCA2)、SMARCA4、骨形态发生蛋白2诱导型激酶(bone morphogenetic protein 2 inducible kinase,BMP2K)、Synergin gamma(SYNRG)、cell division cycle and apoptosis regulator 1(CCAR1)、DUX4、SEC24 homolog B(SEC24B)、泛素特异性蛋白酶25(ubiquitin specific peptidase 25,USP25)、Nipped-B-like protein(NIPBL)、Testis expressed 41(TEX41)、TCF4和网格蛋白重链(clathrin heavy chain,CLTC)[4,6-11]。各融合亚型在AL中的发生频率具有显著差异,EP300-ZNF384的发生率最高。ZNF384融合在ALL中被描述为G5亚型,具有临床相关性,可扰乱B细胞分化并显示JAK-STAT通路的显著激活,因此使用该通路抑制剂治疗应对患者有潜在益处[2,6]。除此外,融合蛋白的主转录活性相较独立的ZNF384发生显著的变化,这些高表达的基因涉及干细胞分化、细胞周期、氧化磷酸化和DNA修复等多种生物学过程[1,2,12-15]。
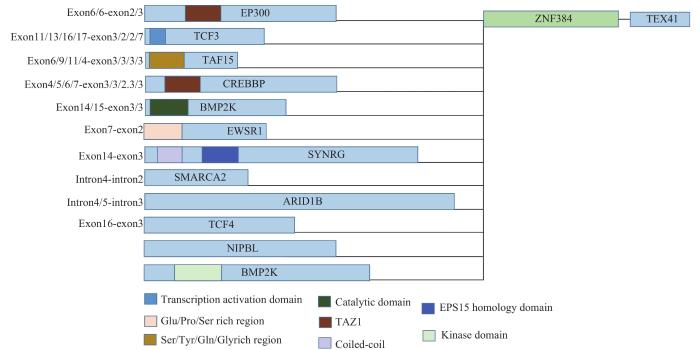
Note: The exon6/6-exon2/3 on the left side of the figure indicates that there are two types of fusion breakpoints between EP300 and ZNF384, exon6-exon2 and exon6-exon3. The small colored squares below the figure represent protein domains. TAZ1—transcriptional adapter zinc binding 1.
Fig 2
Breakpoint statistics and fusion protein structure diagram of some ZNF384 fusion subtypes
FET蛋白与多种转录因子的融合已在多种恶性肿瘤中被发现[57]。TAF15-ZNF384可显著上调Ba/f3细胞中的B、T淋巴细胞衰减因子(B and T lymphocyte attenuator,BTLA)和心肌营养蛋白样细胞因子1(cardiotrophin-like cytokine factor 1,CLCF1)的表达并增强其致癌转化作用,在体外具有转化活性[1,18]。基于TAF15以及EWSR1之间的同源性,研究者认为TAF15-ZNF384和EWSR1-ZNF384之间可能存在类似的导致MPAL的遗传机制[16],但其详细分子机制尚待进一步阐明。除此之外,TAF15-ZNF384病例显示出高度的生物学和临床异质性,TAF15融合的断点位置从外显子4延伸到外显子10、其更广泛的基因突变状态、额外的染色体异常、CD10的表达以及发病年龄段和临床特征的多样性,为这类融合的致癌机制研究带来了更大的挑战[18,48,59]。
TCF3是AL中常见的融合靶点,已报道与TCF3融合的基因有:ZNF384、PBX1、hepatic leukemia factor(HLF)、TCF3 fusion partner(TFPT)[63-65],表达产物除ZNF384外均发生不同程度的缺失。TCF3与ZNF384融合的断点涉及外显子11、13、16和17,保留了AD1和部分AD2结构域[13]。有研究[13]发现TCF3与ZNF384的融合可表达2种融合蛋白(即含6个锌指和8个锌指的ZNF384),这2种融合蛋白的染色体免疫共沉淀测序(chromatin immunoprecipitation followed by sequencing,ChIP-seq)结果显示,6锌指的TCF3-ZNF384即为AL中出现的亚型,该亚型的富集基因参与RNA稳定性的维持和淋巴细胞分化[9]。在TCF3-ZNF384阳性ALL中,基因表达谱与EP300/CREBBP-ZNF384阳性ALL相似,主要影响造血干细胞的分化。最近的一项研究[48]发现ZNF384融合蛋白通过与CTCF和RNA聚合酶Ⅱ相互作用介导染色质环形成,这种基于全基因组染色质的相互作用将FMS样酪氨酸酶3(FMS related receptor tyrosine kinase 3,FLT3)的启动子和增强子区相互连接激活,并增强了FLT3的表达;已知FLT3在所有携带ZNF384融合(EP300、TCF3、TAF15、ARID1B、CLTC、CREBBP、EWSR1、NIPBL和SMARCA2)的病例中始终过表达且常与其他致癌突变同时出现。因此在ZNF384融合的ALL中,似乎存在FLT3信号转导的依赖性,FLT3抑制剂的使用有可能改善患者的临床症状[66-67]。
这些融合伴侣中,除TEX41属于非编码RNA外,其余皆为蛋白编码基因。它们的转录翻译产物大多定位于细胞核。通过分析美国国家生物技术信息中心(National Center of Biotechnology Information,NCBI)以及蛋白质组学数据库(UniProt Protein Resource,UniPort)[68]对这些融合伴侣的功能注释,我们了解到,大多数融合伴侣参与多种细胞过程,尤其是涉及组蛋白乙酰化的染色质重塑过程,常与其他转录因子协作以共调节干细胞分化、神经元的发育和迁移等过程。有趣的是,这些融合伴侣和ZNF384在睾丸和骨髓均高表达,说明ZNF384在选择融合伴侣方面是有规律的,二者之间在生理功能上可能存在一定的联系,融合后则实现了某些功能的强化,打破了原有的调控平衡,使细胞癌变。因此通过研究融合伴侣功能及它们之间的相关性,将对我们研究ZNF384融合致白血病的分子机制带来新的见解。
LI Ying conceived the idea and drafted the original manuscript. MENG Guoyu and Chen li supervised the writing of the article. TAN Yangxia, YIN Hongxin, and JIANG Yanling participated in the reviewing and editing. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
QIAN M X, ZHANG H, KHAM S K, et al. Whole-transcriptome sequencing identifies a distinct subtype of acute lymphoblastic leukemia with predominant genomic abnormalities of EP300 and CREBBP[J]. Genome Res, 2017, 27(2): 185-195.
LI J F, DAI Y T, LILLJEBJÖRN H, et al. Transcriptional landscape of B cell precursor acute lymphoblastic leukemia based on an international study of 1, 223 cases[J]. Proc Natl Acad Sci USA, 2018, 115(50): E11711-E11720.
ZALIOVA M, STUCHLY J, WINKOWSKA L, et al. Genomic landscape of pediatric B-other acute lymphoblastic leukemia in a consecutive European cohort[J]. Haematologica, 2019, 104(7): 1396-1406.
LI J F, DAI Y T, WU L, et al. Emerging molecular subtypes and therapeutic targets in B-cell precursor acute lymphoblastic leukemia[J]. Front Med, 2021, 15(3): 347-371.
MÄKINEN V P, REHN J, BREEN J, et al. Multi-cohort transcriptomic subtyping of B-cell acute lymphoblastic leukemia[J]. Int J Mol Sci, 2022, 23(9): 4574.
QIN Y Z, JIANG Q, XU L P, et al. The prognostic significance of ZNF384 fusions in adult ph-negative B-cell precursor acute lymphoblastic leukemia: a comprehensive cohort study from a single Chinese center[J]. Front Oncol, 2021, 11: 632532.
GOCHO Y, KIYOKAWA N, ICHIKAWA H, et al. A novel recurrent EP300-ZNF384 gene fusion in B-cell precursor acute lymphoblastic leukemia[J]. Leukemia, 2015, 29(12): 2445-2448.
DICKERSON K M, QU C X, GAO Q S, et al. ZNF384 fusion oncoproteins drive lineage aberrancy in acute leukemia[J]. Blood Cancer Discov, 2022, 3(3): 240-263.
WU Z Y, ZHANG F, LIU C Z, et al. Whole transcriptome sequencing reveals a TCF4-ZNF384 fusion in acute lymphoblastic leukemia[J]. Front Oncol, 2022, 12: 900054.
HIRABAYASHI S, BUTLER E R, OHKI K, et al. Clinical characteristics and outcomes of B-ALL with ZNF384 rearrangements: a retrospective analysis by the Ponte di Legno Childhood ALL Working Group[J]. Leukemia, 2021, 35(11): 3272-3277.
ALEXANDER T B, GU Z H, IACOBUCCI I, et al. The genetic basis and cell of origin of mixed phenotype acute leukaemia[J]. Nature, 2018, 562(7727): 373-379.
YAGUCHI A, ISHIBASHI T, TERADA K, et al. EP300-ZNF384 fusion gene product up-regulates GATA3 gene expression and induces hematopoietic stem cell gene expression signature in B-cell precursor acute lymphoblastic leukemia cells[J]. Int J Hematol, 2017, 106(2): 269-281.
MCCLURE B J, HEATLEY S L, KOK C H, et al. Pre-B acute lymphoblastic leukaemia recurrent fusion, EP300-ZNF384, is associated with a distinct gene expression[J]. Br J Cancer, 2018, 118(7): 1000-1004.
YAMAMOTO K, KAWAMOTO S, MIZUTANI Y, et al. Mixed phenotype acute leukemia with t (12;17) (p13;q21)/TAF15-ZNF384 and other chromosome abnormalities[J]. Cytogenet Genome Res, 2016, 149(3): 165-170.
PING N N, QIU H Y, WANG Q, et al. Establishment and genetic characterization of a novel mixed-phenotype acute leukemia cell line with EP300-ZNF384 fusion[J]. J Hematol Oncol, 2015, 8: 100.
MARTINI A, LA STARZA R, JANSSEN H, et al. Recurrent rearrangement of the Ewing′s sarcoma gene, EWSR1, or its homologue, TAF15, with the transcription factor CIZ/NMP4 in acute leukemia[J]. Cancer Res, 2002, 62(19): 5408-5412.
IACOBUCCI I, KIMURA S, MULLIGHAN C G. Biologic and therapeutic implications of genomic alterations in acute lymphoblastic leukemia[J]. J Clin Med, 2021, 10(17): 3792.
MA J, GUAN J, CHEN B. ZNF384 rearrangement in acute lymphocytic leukemia with renal involvement as the first manifestation is associated with a poor prognosis: a case report[J]. Mol Cytogenet, 2022, 15(1): 4.
NISHIMURA A, HASEGAWA D, HIRABAYASHI S, et al. Very late relapse cases of TCF3-ZNF384-positive acute lymphoblastic leukemia[J]. Pediatr Blood Cancer, 2019, 66(11): e27891.
JING Y, LI Y F, WAN H, et al. Detection of EP300-ZNF384 fusion in patients with acute lymphoblastic leukemia using RNA fusion gene panel sequencing[J]. Ann Hematol, 2020, 99(11): 2611-2617.
OBERLEY M J, GAYNON P S, BHOJWANI D, et al. Myeloid lineage switch following chimeric antigen receptor T-cell therapy in a patient with TCF3-ZNF384 fusion-positive B-lymphoblastic leukemia[J]. Pediatr Blood Cancer, 2018, 65(9): e27265.
GERR H, ZIMMERMANN M, SCHRAPPE M, et al. Acute leukaemias of ambiguous lineage in children: characterization, prognosis and therapy recommendations[J]. Br J Haematol, 2010, 149(1): 84-92.
TRAN T H, LANGLOIS S, MELOCHE C, et al. Whole-transcriptome analysis in acute lymphoblastic leukemia: a report from the DFCI ALL Consortium Protocol 16-001[J]. Blood Adv, 2022, 6(4): 1329-1341.
MOORMAN A V, BARRETTA E, BUTLER E R, et al. Prognostic impact of chromosomal abnormalities and copy number alterations in adult B-cell precursor acute lymphoblastic leukaemia: a UKALL14 study[J]. Leukemia, 2022, 36(3): 625-636.
PAIETTA E, ROBERTS K G, WANG V, et al. Molecular classification improves risk assessment in adult BCR-ABL1-negative B-ALL[J]. Blood, 2021, 138(11): 948-958.
JEHA S, CHOI J, ROBERTS K G, et al. Clinical significance of novel subtypes of acute lymphoblastic leukemia in the context of minimal residual disease-directed therapy[J]. Blood Cancer Discov, 2021, 2(4): 326-337.
YAO Z L, LI Y F, LI M, et al. Analysis of clinical characteristics of acute B lymphoblastic leukemia with EP300-ZNF384 fusion gene positive [J]. Journal of Experimental Hematology, 2020, 28(1): 24-28.
SHAGO M, ABLA O, HITZLER J, et al. Frequency and outcome of pediatric acute lymphoblastic leukemia with ZNF384 gene rearrangements including a novel translocation resulting in an ARID1B/ZNF384 gene fusion[J]. Pediatr Blood Cancer, 2016, 63(11): 1915-1921.
FAN Z Y, TARDIF G, BOILEAU C, et al. Identification in human osteoarthritic chondrocytes of proteins binding to the novel regulatory site AGRE in the human matrix metalloprotease 13 proximal promoter[J]. Arthritis Rheum, 2006, 54(8): 2471-2480.
YOUNG S K, SHAO Y, BIDWELL J P, et al. Nuclear matrix protein 4 is a novel regulator of ribosome biogenesis and controls the unfolded protein response via repression of Gadd34 expression[J]. J Biol Chem, 2016, 291(26): 13780-13788.
JIN H L, VAN'T HOF R J, ALBAGHA O M, et al. Promoter and intron 1 polymorphisms of COL1A1 interact to regulate transcription and susceptibility to osteoporosis[J]. Hum Mol Genet, 2009, 18(15): 2729-2738.
SHAO Y, WICHERN E, CHILDRESS P J, et al. Loss of Nmp4 optimizes osteogenic metabolism and secretion to enhance bone quality[J]. Am J Physiol Endocrinol Metab, 2019, 316(5): E749-E772.
GAO Y Y, LING Z Y, ZHU Y R, et al. The histone acetyltransferase HBO1 functions as a novel oncogenic gene in osteosarcoma[J]. Theranostics, 2021, 11(10): 4599-4615.
WAN F, ZHOU J, CHEN X, et al. Overexpression and mutation of ZNF384 is associated with favorable prognosis in breast cancer patients[J]. Transl Cancer Res, 2019, 8(3): 779-787.
MENG Q X, WANG K N, LI J H, et al. ZNF384-ZEB1 feedback loop regulates breast cancer metastasis[J]. Mol Med, 2022, 28(1): 111.
SINGH J K, SMITH R, ROTHER M B, et al. Zinc finger protein ZNF384 is an adaptor of Ku to DNA during classical non-homologous end-joining[J]. Nat Commun, 2021, 12(1): 6560.
CHILDRESS P, STAYROOK K R, ALVAREZ M B, et al. Genome-wide mapping and interrogation of the Nmp4 antianabolic bone axis[J]. Mol Endocrinol, 2015, 29(9): 1269-1285.
LIU S G, YUAN X Q, SU H, et al. ZNF384: a potential therapeutic target for psoriasis and alzheimer's disease through inflammation and metabolism[J]. Front Immunol, 2022, 13: 892368.
NYQUIST K B, THORSEN J, ZELLER B, et al. Identification of the TAF15-ZNF384 fusion gene in two new cases of acute lymphoblastic leukemia with a t (12;17) (p13;q12)[J]. Cancer Genet, 2011, 204(3): 147-152.
BLACK J C, CHOI J E, LOMBARDO S R, et al. A mechanism for coordinating chromatin modification and preinitiation complex assembly[J]. Mol Cell, 2006, 23(6): 809-818.
ZHANG X, YUAN X, ZHU W, et al. SALL4: an emerging cancer biomarker and target[J]. Cancer Lett, 2015, 357(1): 55-62.
JIANG Y, JI Q K, LONG X Y, et al. CLCF1 is a novel potential immune-related target with predictive value for prognosis and immunotherapy response in glioma[J]. Front Immunol, 2022, 13: 810832.
DEMERLÉ C, GORVEL L, OLIVE D. BTLA-HVEM couple in health and diseases: insights for immunotherapy in lung cancer[J]. Front Oncol, 2021, 11: 682007.
ALVES J, WURDAK H, GARAY-MALPARTIDA H M, et al. TAF15 and the leukemia-associated fusion protein TAF15-CIZ/NMP4 are cleaved by caspases-3 and-7[J]. Biochem Biophys Res Commun, 2009, 384(4): 495-500.
GEORGAKOPOULOS N, DIAMANTOPOULOS P, MICCI F, et al. An adult patient with early pre-B acute lymphoblastic leukemia with t(12;17)(p13;q21)/ZNF384-TAF15[J]. In Vivo, 2018, 32(5): 1241-1245.
LIANG J J, PENG H, WANG J J, et al. Relationship between the structure and function of the transcriptional regulator E2A[J]. J Biol Res (Thessalon), 2021, 28(1): 15.
RAO C, MALAGUTI M, MASON J O, et al. The transcription factor E2A drives neural differentiation in pluripotent cells[J]. Development, 2020, 147(12): dev184093.
LÓPEZ-MENÉNDEZ C, VÁZQUEZ-NAHARRO A, SANTOS V, et al. E2A modulates stemness, metastasis, and therapeutic resistance of breast cancer[J]. Cancer Res, 2021, 81(17): 4529-4544.
ZHOU B Q, CHU X R, TIAN H, et al. The clinical outcomes and genomic landscapes of acute lymphoblastic leukemia patients with E2A-PBX1: a 10-year retrospective study[J]. Am J Hematol, 2021, 96(11): 1461-1471.
BRAMBILLASCA F, MOSNA G, BALLABIO E, et al. Promoter analysis of TFPT (FB1), a molecular partner of TCF3 (E2A) in childhood acute lymphoblastic leukemia[J]. Biochem Biophys Res Commun, 2001, 288(5): 1250-1257.
BAUDIS M, PRIMA V, TUNG Y H, et al. ABCB1 over-expression and drug-efflux in acute lymphoblastic leukemia cell lines with t(17;19) and E2A-HLF expression[J]. Pediatr Blood Cancer, 2006, 47(6): 757-764.
ZHAO X J, WANG P, DIEDRICH J D, et al. Epigenetic activation of the FLT3 gene by ZNF384 fusion confers a therapeutic susceptibility in acute lymphoblastic leukemia[J]. Nat Commun, 2022, 13(1): 5401.
GRIFFITH M, GRIFFITH O L, KRYSIAK K, et al. Comprehensive genomic analysis reveals FLT3 activation and a therapeutic strategy for a patient with relapsed adult B-lymphoblastic leukemia[J]. Exp Hematol, 2016, 44(7): 603-613.
... FET蛋白与多种转录因子的融合已在多种恶性肿瘤中被发现[57].TAF15-ZNF384可显著上调Ba/f3细胞中的B、T淋巴细胞衰减因子(B and T lymphocyte attenuator,BTLA)和心肌营养蛋白样细胞因子1(cardiotrophin-like cytokine factor 1,CLCF1)的表达并增强其致癌转化作用,在体外具有转化活性[1,18].基于TAF15以及EWSR1之间的同源性,研究者认为TAF15-ZNF384和EWSR1-ZNF384之间可能存在类似的导致MPAL的遗传机制[16],但其详细分子机制尚待进一步阐明.除此之外,TAF15-ZNF384病例显示出高度的生物学和临床异质性,TAF15融合的断点位置从外显子4延伸到外显子10、其更广泛的基因突变状态、额外的染色体异常、CD10的表达以及发病年龄段和临床特征的多样性,为这类融合的致癌机制研究带来了更大的挑战[18,48,59]. ...
6
... 急性白血病(acute leukemia,AL)是造血干细胞恶性增殖的克隆性疾病,包括急性淋巴细胞白血病(acute lymphoblastic leukemia,ALL)、急性髓系白血病(acute myeloid leukemia,AML)、混合表型急性白血病(mixed phenotype acute leukemia,MPAL).染色体易位已成为许多血液系统恶性肿瘤的基因组标志,异常融合基因编码的产物主要与造血分化、细胞增殖和细胞周期等机制的转录因子有关[1].二代测序揭示了AL中存在的多种染色体易位,如肌细胞增强因子2D(myocyte enhancer factor 2D,MEF2D)融合、T细胞因子3(T cell factor 3,TCF3)-前B淋巴细胞白血病转录因子(pre-B-cell leukemia transcription factor1,PBX1)、ETS variant transcription factor 6(ETV6)-RUNX家族相关转录因子1(RUNX family transcription factor 1,RUNX1)阳性/ETV6-RUNX1样、Double homeobox 4(DUX4)融合、ZNF384融合、BCR-ABL1/Ph样和Lysine methyltransferase 2A (KMT2A)融合相关等[1-7].其中ZNF384融合的存在较为特殊,迄今为止,已报道了18个与ZNF384发生融合的伴侣基因:E1A结合蛋白p300(E1A binding protein p300,EP300)、转录因子3(transcription factor 3,TCF3)、TATA-box binding protein associated factor 15(TAF15)、cAMP反应元件结合蛋白(cAMP-response element-binding protein,CREB)结合蛋白(CREB-binding protein,CREBBP)、EWS RNA binding protein 1(EWSR1)、转录因子AT富集区1B(AT-rich interaction domain 1B,ARID1B)、SWI/SNF related,matrix associated,actin dependent regulator of chromatin,subfamily a,member 2(SMARCA2)、SMARCA4、骨形态发生蛋白2诱导型激酶(bone morphogenetic protein 2 inducible kinase,BMP2K)、Synergin gamma(SYNRG)、cell division cycle and apoptosis regulator 1(CCAR1)、DUX4、SEC24 homolog B(SEC24B)、泛素特异性蛋白酶25(ubiquitin specific peptidase 25,USP25)、Nipped-B-like protein(NIPBL)、Testis expressed 41(TEX41)、TCF4和网格蛋白重链(clathrin heavy chain,CLTC)[4,6-11].各融合亚型在AL中的发生频率具有显著差异,EP300-ZNF384的发生率最高.ZNF384融合在ALL中被描述为G5亚型,具有临床相关性,可扰乱B细胞分化并显示JAK-STAT通路的显著激活,因此使用该通路抑制剂治疗应对患者有潜在益处[2,6].除此外,融合蛋白的主转录活性相较独立的ZNF384发生显著的变化,这些高表达的基因涉及干细胞分化、细胞周期、氧化磷酸化和DNA修复等多种生物学过程[1,2,12-15]. ...
... TCF3是AL中常见的融合靶点,已报道与TCF3融合的基因有:ZNF384、PBX1、hepatic leukemia factor(HLF)、TCF3 fusion partner(TFPT)[63-65],表达产物除ZNF384外均发生不同程度的缺失.TCF3与ZNF384融合的断点涉及外显子11、13、16和17,保留了AD1和部分AD2结构域[13].有研究[13]发现TCF3与ZNF384的融合可表达2种融合蛋白(即含6个锌指和8个锌指的ZNF384),这2种融合蛋白的染色体免疫共沉淀测序(chromatin immunoprecipitation followed by sequencing,ChIP-seq)结果显示,6锌指的TCF3-ZNF384即为AL中出现的亚型,该亚型的富集基因参与RNA稳定性的维持和淋巴细胞分化[9].在TCF3-ZNF384阳性ALL中,基因表达谱与EP300/CREBBP-ZNF384阳性ALL相似,主要影响造血干细胞的分化.最近的一项研究[48]发现ZNF384融合蛋白通过与CTCF和RNA聚合酶Ⅱ相互作用介导染色质环形成,这种基于全基因组染色质的相互作用将FMS样酪氨酸酶3(FMS related receptor tyrosine kinase 3,FLT3)的启动子和增强子区相互连接激活,并增强了FLT3的表达;已知FLT3在所有携带ZNF384融合(EP300、TCF3、TAF15、ARID1B、CLTC、CREBBP、EWSR1、NIPBL和SMARCA2)的病例中始终过表达且常与其他致癌突变同时出现.因此在ZNF384融合的ALL中,似乎存在FLT3信号转导的依赖性,FLT3抑制剂的使用有可能改善患者的临床症状[66-67]. ...
... [13]发现TCF3与ZNF384的融合可表达2种融合蛋白(即含6个锌指和8个锌指的ZNF384),这2种融合蛋白的染色体免疫共沉淀测序(chromatin immunoprecipitation followed by sequencing,ChIP-seq)结果显示,6锌指的TCF3-ZNF384即为AL中出现的亚型,该亚型的富集基因参与RNA稳定性的维持和淋巴细胞分化[9].在TCF3-ZNF384阳性ALL中,基因表达谱与EP300/CREBBP-ZNF384阳性ALL相似,主要影响造血干细胞的分化.最近的一项研究[48]发现ZNF384融合蛋白通过与CTCF和RNA聚合酶Ⅱ相互作用介导染色质环形成,这种基于全基因组染色质的相互作用将FMS样酪氨酸酶3(FMS related receptor tyrosine kinase 3,FLT3)的启动子和增强子区相互连接激活,并增强了FLT3的表达;已知FLT3在所有携带ZNF384融合(EP300、TCF3、TAF15、ARID1B、CLTC、CREBBP、EWSR1、NIPBL和SMARCA2)的病例中始终过表达且常与其他致癌突变同时出现.因此在ZNF384融合的ALL中,似乎存在FLT3信号转导的依赖性,FLT3抑制剂的使用有可能改善患者的临床症状[66-67]. ...
... FET蛋白与多种转录因子的融合已在多种恶性肿瘤中被发现[57].TAF15-ZNF384可显著上调Ba/f3细胞中的B、T淋巴细胞衰减因子(B and T lymphocyte attenuator,BTLA)和心肌营养蛋白样细胞因子1(cardiotrophin-like cytokine factor 1,CLCF1)的表达并增强其致癌转化作用,在体外具有转化活性[1,18].基于TAF15以及EWSR1之间的同源性,研究者认为TAF15-ZNF384和EWSR1-ZNF384之间可能存在类似的导致MPAL的遗传机制[16],但其详细分子机制尚待进一步阐明.除此之外,TAF15-ZNF384病例显示出高度的生物学和临床异质性,TAF15融合的断点位置从外显子4延伸到外显子10、其更广泛的基因突变状态、额外的染色体异常、CD10的表达以及发病年龄段和临床特征的多样性,为这类融合的致癌机制研究带来了更大的挑战[18,48,59]. ...
... FET蛋白与多种转录因子的融合已在多种恶性肿瘤中被发现[57].TAF15-ZNF384可显著上调Ba/f3细胞中的B、T淋巴细胞衰减因子(B and T lymphocyte attenuator,BTLA)和心肌营养蛋白样细胞因子1(cardiotrophin-like cytokine factor 1,CLCF1)的表达并增强其致癌转化作用,在体外具有转化活性[1,18].基于TAF15以及EWSR1之间的同源性,研究者认为TAF15-ZNF384和EWSR1-ZNF384之间可能存在类似的导致MPAL的遗传机制[16],但其详细分子机制尚待进一步阐明.除此之外,TAF15-ZNF384病例显示出高度的生物学和临床异质性,TAF15融合的断点位置从外显子4延伸到外显子10、其更广泛的基因突变状态、额外的染色体异常、CD10的表达以及发病年龄段和临床特征的多样性,为这类融合的致癌机制研究带来了更大的挑战[18,48,59]. ...
... FET蛋白与多种转录因子的融合已在多种恶性肿瘤中被发现[57].TAF15-ZNF384可显著上调Ba/f3细胞中的B、T淋巴细胞衰减因子(B and T lymphocyte attenuator,BTLA)和心肌营养蛋白样细胞因子1(cardiotrophin-like cytokine factor 1,CLCF1)的表达并增强其致癌转化作用,在体外具有转化活性[1,18].基于TAF15以及EWSR1之间的同源性,研究者认为TAF15-ZNF384和EWSR1-ZNF384之间可能存在类似的导致MPAL的遗传机制[16],但其详细分子机制尚待进一步阐明.除此之外,TAF15-ZNF384病例显示出高度的生物学和临床异质性,TAF15融合的断点位置从外显子4延伸到外显子10、其更广泛的基因突变状态、额外的染色体异常、CD10的表达以及发病年龄段和临床特征的多样性,为这类融合的致癌机制研究带来了更大的挑战[18,48,59]. ...
... TCF3是AL中常见的融合靶点,已报道与TCF3融合的基因有:ZNF384、PBX1、hepatic leukemia factor(HLF)、TCF3 fusion partner(TFPT)[63-65],表达产物除ZNF384外均发生不同程度的缺失.TCF3与ZNF384融合的断点涉及外显子11、13、16和17,保留了AD1和部分AD2结构域[13].有研究[13]发现TCF3与ZNF384的融合可表达2种融合蛋白(即含6个锌指和8个锌指的ZNF384),这2种融合蛋白的染色体免疫共沉淀测序(chromatin immunoprecipitation followed by sequencing,ChIP-seq)结果显示,6锌指的TCF3-ZNF384即为AL中出现的亚型,该亚型的富集基因参与RNA稳定性的维持和淋巴细胞分化[9].在TCF3-ZNF384阳性ALL中,基因表达谱与EP300/CREBBP-ZNF384阳性ALL相似,主要影响造血干细胞的分化.最近的一项研究[48]发现ZNF384融合蛋白通过与CTCF和RNA聚合酶Ⅱ相互作用介导染色质环形成,这种基于全基因组染色质的相互作用将FMS样酪氨酸酶3(FMS related receptor tyrosine kinase 3,FLT3)的启动子和增强子区相互连接激活,并增强了FLT3的表达;已知FLT3在所有携带ZNF384融合(EP300、TCF3、TAF15、ARID1B、CLTC、CREBBP、EWSR1、NIPBL和SMARCA2)的病例中始终过表达且常与其他致癌突变同时出现.因此在ZNF384融合的ALL中,似乎存在FLT3信号转导的依赖性,FLT3抑制剂的使用有可能改善患者的临床症状[66-67]. ...
... FET蛋白与多种转录因子的融合已在多种恶性肿瘤中被发现[57].TAF15-ZNF384可显著上调Ba/f3细胞中的B、T淋巴细胞衰减因子(B and T lymphocyte attenuator,BTLA)和心肌营养蛋白样细胞因子1(cardiotrophin-like cytokine factor 1,CLCF1)的表达并增强其致癌转化作用,在体外具有转化活性[1,18].基于TAF15以及EWSR1之间的同源性,研究者认为TAF15-ZNF384和EWSR1-ZNF384之间可能存在类似的导致MPAL的遗传机制[16],但其详细分子机制尚待进一步阐明.除此之外,TAF15-ZNF384病例显示出高度的生物学和临床异质性,TAF15融合的断点位置从外显子4延伸到外显子10、其更广泛的基因突变状态、额外的染色体异常、CD10的表达以及发病年龄段和临床特征的多样性,为这类融合的致癌机制研究带来了更大的挑战[18,48,59]. ...
... FET蛋白与多种转录因子的融合已在多种恶性肿瘤中被发现[57].TAF15-ZNF384可显著上调Ba/f3细胞中的B、T淋巴细胞衰减因子(B and T lymphocyte attenuator,BTLA)和心肌营养蛋白样细胞因子1(cardiotrophin-like cytokine factor 1,CLCF1)的表达并增强其致癌转化作用,在体外具有转化活性[1,18].基于TAF15以及EWSR1之间的同源性,研究者认为TAF15-ZNF384和EWSR1-ZNF384之间可能存在类似的导致MPAL的遗传机制[16],但其详细分子机制尚待进一步阐明.除此之外,TAF15-ZNF384病例显示出高度的生物学和临床异质性,TAF15融合的断点位置从外显子4延伸到外显子10、其更广泛的基因突变状态、额外的染色体异常、CD10的表达以及发病年龄段和临床特征的多样性,为这类融合的致癌机制研究带来了更大的挑战[18,48,59]. ...
... TCF3是AL中常见的融合靶点,已报道与TCF3融合的基因有:ZNF384、PBX1、hepatic leukemia factor(HLF)、TCF3 fusion partner(TFPT)[63-65],表达产物除ZNF384外均发生不同程度的缺失.TCF3与ZNF384融合的断点涉及外显子11、13、16和17,保留了AD1和部分AD2结构域[13].有研究[13]发现TCF3与ZNF384的融合可表达2种融合蛋白(即含6个锌指和8个锌指的ZNF384),这2种融合蛋白的染色体免疫共沉淀测序(chromatin immunoprecipitation followed by sequencing,ChIP-seq)结果显示,6锌指的TCF3-ZNF384即为AL中出现的亚型,该亚型的富集基因参与RNA稳定性的维持和淋巴细胞分化[9].在TCF3-ZNF384阳性ALL中,基因表达谱与EP300/CREBBP-ZNF384阳性ALL相似,主要影响造血干细胞的分化.最近的一项研究[48]发现ZNF384融合蛋白通过与CTCF和RNA聚合酶Ⅱ相互作用介导染色质环形成,这种基于全基因组染色质的相互作用将FMS样酪氨酸酶3(FMS related receptor tyrosine kinase 3,FLT3)的启动子和增强子区相互连接激活,并增强了FLT3的表达;已知FLT3在所有携带ZNF384融合(EP300、TCF3、TAF15、ARID1B、CLTC、CREBBP、EWSR1、NIPBL和SMARCA2)的病例中始终过表达且常与其他致癌突变同时出现.因此在ZNF384融合的ALL中,似乎存在FLT3信号转导的依赖性,FLT3抑制剂的使用有可能改善患者的临床症状[66-67]. ...
0
1
... TCF3是AL中常见的融合靶点,已报道与TCF3融合的基因有:ZNF384、PBX1、hepatic leukemia factor(HLF)、TCF3 fusion partner(TFPT)[63-65],表达产物除ZNF384外均发生不同程度的缺失.TCF3与ZNF384融合的断点涉及外显子11、13、16和17,保留了AD1和部分AD2结构域[13].有研究[13]发现TCF3与ZNF384的融合可表达2种融合蛋白(即含6个锌指和8个锌指的ZNF384),这2种融合蛋白的染色体免疫共沉淀测序(chromatin immunoprecipitation followed by sequencing,ChIP-seq)结果显示,6锌指的TCF3-ZNF384即为AL中出现的亚型,该亚型的富集基因参与RNA稳定性的维持和淋巴细胞分化[9].在TCF3-ZNF384阳性ALL中,基因表达谱与EP300/CREBBP-ZNF384阳性ALL相似,主要影响造血干细胞的分化.最近的一项研究[48]发现ZNF384融合蛋白通过与CTCF和RNA聚合酶Ⅱ相互作用介导染色质环形成,这种基于全基因组染色质的相互作用将FMS样酪氨酸酶3(FMS related receptor tyrosine kinase 3,FLT3)的启动子和增强子区相互连接激活,并增强了FLT3的表达;已知FLT3在所有携带ZNF384融合(EP300、TCF3、TAF15、ARID1B、CLTC、CREBBP、EWSR1、NIPBL和SMARCA2)的病例中始终过表达且常与其他致癌突变同时出现.因此在ZNF384融合的ALL中,似乎存在FLT3信号转导的依赖性,FLT3抑制剂的使用有可能改善患者的临床症状[66-67]. ...
1
... TCF3是AL中常见的融合靶点,已报道与TCF3融合的基因有:ZNF384、PBX1、hepatic leukemia factor(HLF)、TCF3 fusion partner(TFPT)[63-65],表达产物除ZNF384外均发生不同程度的缺失.TCF3与ZNF384融合的断点涉及外显子11、13、16和17,保留了AD1和部分AD2结构域[13].有研究[13]发现TCF3与ZNF384的融合可表达2种融合蛋白(即含6个锌指和8个锌指的ZNF384),这2种融合蛋白的染色体免疫共沉淀测序(chromatin immunoprecipitation followed by sequencing,ChIP-seq)结果显示,6锌指的TCF3-ZNF384即为AL中出现的亚型,该亚型的富集基因参与RNA稳定性的维持和淋巴细胞分化[9].在TCF3-ZNF384阳性ALL中,基因表达谱与EP300/CREBBP-ZNF384阳性ALL相似,主要影响造血干细胞的分化.最近的一项研究[48]发现ZNF384融合蛋白通过与CTCF和RNA聚合酶Ⅱ相互作用介导染色质环形成,这种基于全基因组染色质的相互作用将FMS样酪氨酸酶3(FMS related receptor tyrosine kinase 3,FLT3)的启动子和增强子区相互连接激活,并增强了FLT3的表达;已知FLT3在所有携带ZNF384融合(EP300、TCF3、TAF15、ARID1B、CLTC、CREBBP、EWSR1、NIPBL和SMARCA2)的病例中始终过表达且常与其他致癌突变同时出现.因此在ZNF384融合的ALL中,似乎存在FLT3信号转导的依赖性,FLT3抑制剂的使用有可能改善患者的临床症状[66-67]. ...
1
... TCF3是AL中常见的融合靶点,已报道与TCF3融合的基因有:ZNF384、PBX1、hepatic leukemia factor(HLF)、TCF3 fusion partner(TFPT)[63-65],表达产物除ZNF384外均发生不同程度的缺失.TCF3与ZNF384融合的断点涉及外显子11、13、16和17,保留了AD1和部分AD2结构域[13].有研究[13]发现TCF3与ZNF384的融合可表达2种融合蛋白(即含6个锌指和8个锌指的ZNF384),这2种融合蛋白的染色体免疫共沉淀测序(chromatin immunoprecipitation followed by sequencing,ChIP-seq)结果显示,6锌指的TCF3-ZNF384即为AL中出现的亚型,该亚型的富集基因参与RNA稳定性的维持和淋巴细胞分化[9].在TCF3-ZNF384阳性ALL中,基因表达谱与EP300/CREBBP-ZNF384阳性ALL相似,主要影响造血干细胞的分化.最近的一项研究[48]发现ZNF384融合蛋白通过与CTCF和RNA聚合酶Ⅱ相互作用介导染色质环形成,这种基于全基因组染色质的相互作用将FMS样酪氨酸酶3(FMS related receptor tyrosine kinase 3,FLT3)的启动子和增强子区相互连接激活,并增强了FLT3的表达;已知FLT3在所有携带ZNF384融合(EP300、TCF3、TAF15、ARID1B、CLTC、CREBBP、EWSR1、NIPBL和SMARCA2)的病例中始终过表达且常与其他致癌突变同时出现.因此在ZNF384融合的ALL中,似乎存在FLT3信号转导的依赖性,FLT3抑制剂的使用有可能改善患者的临床症状[66-67]. ...
1
... 这些融合伴侣中,除TEX41属于非编码RNA外,其余皆为蛋白编码基因.它们的转录翻译产物大多定位于细胞核.通过分析美国国家生物技术信息中心(National Center of Biotechnology Information,NCBI)以及蛋白质组学数据库(UniProt Protein Resource,UniPort)[68]对这些融合伴侣的功能注释,我们了解到,大多数融合伴侣参与多种细胞过程,尤其是涉及组蛋白乙酰化的染色质重塑过程,常与其他转录因子协作以共调节干细胞分化、神经元的发育和迁移等过程.有趣的是,这些融合伴侣和ZNF384在睾丸和骨髓均高表达,说明ZNF384在选择融合伴侣方面是有规律的,二者之间在生理功能上可能存在一定的联系,融合后则实现了某些功能的强化,打破了原有的调控平衡,使细胞癌变.因此通过研究融合伴侣功能及它们之间的相关性,将对我们研究ZNF384融合致白血病的分子机制带来新的见解. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}