Screening of AAK1 interaction proteins and its role in regulating global translation level in cells
JIANG Guixian,1,2,3, HU Ronggui4, WU Hao,1,2,3
1.Department of Otolaryngology-Head and Neck Surgery, Shanghai Ninth People′s Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai 200011, China
2.Ear Institute, Shanghai Jiao Tong University School of Medicine, Shanghai 200092, China
3.Shanghai Key Laboratory of Translation Medicine on Ear and Nose Diseases, Shanghai 200092, China
4.Center for Excellence in Molecular Cell Science, University of Chinese Academy of Sciences, Shanghai 200032, China
目的·探究衔接子相关蛋白激酶1(adaptor-associated protein kinase 1,AAK1)新的相互作用蛋白,以及除网格蛋白介导的内吞作用外AAK1介导的生物学功能。方法·通过在HEK-293T细胞中分别外源性转染带有标签的AAK1载体与空白对照载体,利用标签特异性的琼脂糖凝胶进行免疫共沉淀(co-immunoprecipitation,CoIP),并联合质谱分析的方法获得潜在与AAK1相互作用的蛋白;通过CoIP初步验证质谱结果;通过荧光共聚焦成像观察AAK1与其潜在结合蛋白在细胞内的空间定位;通过体外纯化重组蛋白,利用谷胱甘肽巯基转移酶融合蛋白沉降实验(glutathione-S-transferase pulldown,GST Pulldown)进一步明确蛋白间是否为直接的相互作用;通过嘌呤霉素结合实验观察AAK1对于细胞内整体翻译水平的调控作用。结果·质谱结果提示AAK1可能与以脆性X相关蛋白1(fragile X mental retardation syndrome-related protein 1,FXR1)、FXR2、脆性X智力低下蛋白(fragile X mental retardation protein 1,FMRP)三者为核心的一系列蛋白形成复合体。外源性转染AAK1-3xFLAG及FMRP-MYC质粒,利用抗FLAG琼脂糖凝胶富集AAK1-3xFLAG后,可以检测到FMRP-MYC的表达;利用内源性抗体进行CoIP,发现富集AAK1可以检测到FMRP的表达。荧光共聚焦成像显示EGFP-AAK1与mCherry-FMRP在细胞质中存在部分空间共定位。GST Pulldown显示FMRP可以直接沉淀HIS6-AAK1重组蛋白。嘌呤霉素结合实验显示相同时间内嘌呤霉素标记的细胞内新合成肽段数量与AAK1蛋白表达量呈正相关。结论·AAK1与FMRP在细胞质内存在直接的相互作用,且AAK1可以提高细胞内的翻译水平。
关键词:衔接子相关蛋白激酶1
;
翻译
;
质谱
;
蛋白相互作用
Abstract
Objective ·To investigate noval interacting partners for adaptor-associated protein kinase 1 (AAK1) and AAK1-mediated biological functions besides clathrin-mediated endocytosis. Methods ·The labeled AAK1 vector and the blank control vector were transfected in HEK-293T cells, and the potential AAK1 interacting proteins were obtained by co-immunoprecipitation with agar-specific gel and mass spectrometry. Further verifications were performed by CoIP and fluorescence-based imaging. Recombinant proteins were purified in vitro and the direct interaction between proteins were confirmed by glutathione-S-transferase pulldown (GST Pulldown) assay. The regulation of AAK1 in the global protein synthesis was explored by puromycin incorporation assay. Results ·Mass spectrometry results showed that AAK1 was associated with a series of proteins, including fragile X mental retardation syndrome-related protein 1 (FXR1), FXR2 and fragile X mental retardation protein 1 (FMRP). Enriching with anti-FLAG agarose gels after exogenous transfecting of AAK1-3xFLAG and FMRP-MYC plasmids, the expression of FMRP-MYC was detected. The expression of FMRP could also be detected by CoIP with endogenous AAK1 antibodies. Fluorescence-based imaging showed that they were spatially colocalized in the cytoplasm. GST Pulldown assay showed that FMRP could pulldown recombinant HIS6-AAK1 protein. Puromycin incorporation assay showed that in the same amount of time, the number of newly synthesized peptides labeled with puromycin was positively correlated with AAK1 protein expression. Conclusion ·AAK1 directly interacts with FMRP in cytoplasm and could up-regulate global protein synthesis level.
Keywords:adaptor-associated protein kinase 1 (AAK1)
;
translation
;
mass spectroscopy
;
protein-protein interaction
JIANG Guixian, HU Ronggui, WU Hao. Screening of AAK1 interaction proteins and its role in regulating global translation level in cells. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(7): 821-828 doi:10.3969/j.issn.1674-8115.2023.07.004
衔接子相关蛋白激酶1(adaptor-associated protein kinase 1,AAK1)于2002年被首次鉴定,并发现其与网格蛋白介导的内吞作用(clathrin-mediated endocytosis,CME)中的关键分子衔接蛋白2(adaptor protein2,AP2)结合,通过磷酸化AP2的μ亚基(AP2 complex subunit μ,AP2M1)对CME过程进行调控[1-2]。因此,AAK1也被称为AP2相关蛋白激酶1。目前,关于AAK1的生物学功能主要围绕CME展开。研究发现,在Wnt信号刺激下,AAK1通过加强AP2M1磷酸化程度,使低密度脂蛋白相关受体6(low-density lipoprotein receptor related protein 6,LRP6)通过CME途径从细胞膜转移到细胞质内[3];AAK1磷酸化内吞衔接蛋白Numb,使其从细胞膜通过CME途径进入内体[4]。除了磷酸化作用外,AAK1还以非磷酸化方式直接结合并稳定活化形式的神经源性基因座缺口同源蛋白1(neurogenic locus notch homolog protein 1,Notch1),使其进入内体并增强其转录活性[5]。AAK1也参与疾病的发生和发展。超氧化物歧化酶1蛋白(superoxide dismutase 1,SOD1)突变(SOD1G85R)与肌萎缩侧索硬化有关,而AAK1仅与SOD1G85R结合且不与SOD1WT结合,并且AAK1蛋白水平在肌萎缩侧索硬化患者中较正常人低[6]。部分病毒通过CME途径进入人体细胞,而AAK1参与调节丙型肝炎病毒[7]、登革热病毒[8]、狂犬病病毒[9]以及在全球范围内流行的新型冠状病毒[10]的进入与细胞间的扩散。因此,也有针对AAK1开展的各类化合物的研发[11-12]。鉴于AAK1对生理、病理过程均有广泛的调控作用,有必要对AAK1开展更深入的研究,探索其更多的分子机制。
脆性X智力低下蛋白(fragile X mental retardation protein 1,FMRP)是脆性X相关(fragile X mental retardation syndrome-related,FXR)蛋白家族的一员,其与脆性X相关蛋白1(FXR protein 1,FXR1)和FXR2具有显著的同源性,在结构、功能上相似[13]。FXR蛋白家族的表达异常是脆性X综合征的直接病因。该病是以智力低下和自闭症样行为为主的一类遗传性疾病,其中以FMRP异常为主[14]。FMRP的功能繁多,可以从多个层面对机体进行调控:① 在DNA层面,调控转录因子与染色质修饰酶的表达与活性[15-16]。② 在蛋白层面,通过直接与离子通道蛋白结合进而影响其兴奋性[17]。③ 在RNA层面,由于FMRP是RNA结合蛋白,FMRP还通过诱导核糖体停滞[18]、与RNA直接结合调控其稳定性[19]、调控RNA在细胞内定位[20]等方式参与调控RNA代谢。运用定量质谱比较青春期和成年期的Fmrp敲除和野生型2种小鼠皮层突触的蛋白质表达情况发现,Fmrp敲除使青春期小鼠大量突触蛋白上调,而在成年小鼠中Fmrp敲除影响较小,表明FMRP的调控还具有时间特异性[21]。由此可见,FMRP的分子机制是十分复杂的。目前,对FMRP的认识相较于FXR1、FXR2更加深入,研究上常以对FMRP的研究为基础反推至FXR1与FXR2。
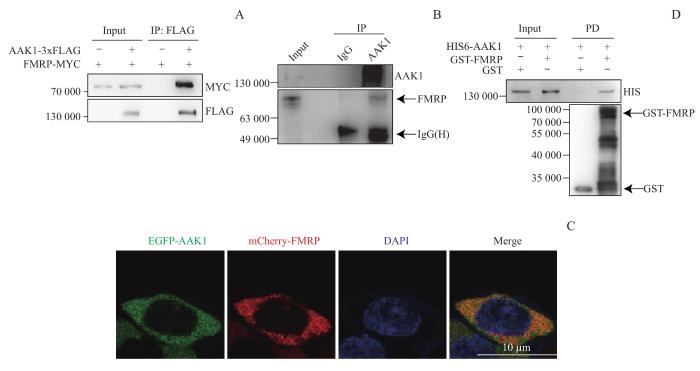
运用CoIP联合质谱(CoIP/MS)分析的方法(图1A),共获得了551个潜在与AAK1结合的候选蛋白。对CoIP过程中留取的部分样品进行考马斯亮蓝染色,发现实验组在AAK1预期大小的140 000处存在清晰条带,而空白对照组未在该位置观察到明显条带,且Western blotting结果表明该条带特异性表达FLAG标签(图1B)。以上结果表明,实验成功富集到了AAK1。对质谱结果进行整理并罗列其中部分结果,发现AAK1表达量在实验组中最多,而对照组中未被鉴定到;并且很多已知与AAK1结合的蛋白,如AP2复合物各个亚基和重肽网格蛋白(clathrin heavy chain,CLTC)等均被鉴定到,提示质谱结果具有一定的可信度和参考价值。此外,在该AAK1相互作用蛋白质谱中还鉴定到一部分以FXR蛋白家族为核心的相互作用的蛋白(图2)。
Note: Poteintial AAK1 interactors were divided into AAK1-associated proteins and FMRP-associated proteins. Red, yellow and green respectively represents high, medium and low level of relative protein expression. AAK1, FMRP, FXR1 and FXR2 are highlighted in red.
Note: A.AAK1-3xFLAG and FMRP-MYC formed a complex in HEK-293T cells, detected by CoIP assay by using anti-FLAG beads, followed by immunoblotting with anti-MYC and anti-FLAG antibodies. B. Endogenous AAK1 and FMRP formed a complex in HEK-293T cells, as demonstrated by CoIP assay by using anti-IgG or anti-AAK1, followed by immunoblotting with anti-FMRP or anti-AAK1 antibodies. C. EGFP-tagged AAK1 co-localized with mCherry-tagged FMRP in cytoplasm of Hela cells. Hela cells were co-transfected with plasmids encoding mCherry-FMRP and EGFP-AAK1. The nuclear of cells was visualized with DAPI staining, and then the fluorescence were detected by confocal microscopy. D. GST Pulldown assays indicated that FMRP protein interacted directly with HIS6-tagged AAK1. PD—GST Pulldown.
Fig 3
Direct interaction between AAK1 and FMRP in cytoplasm
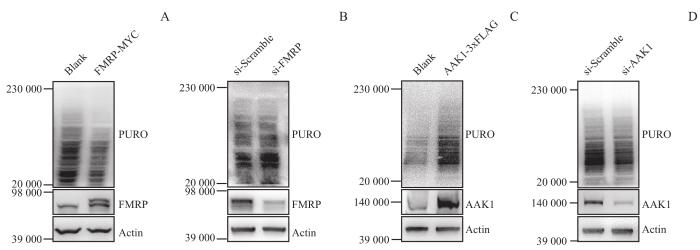
Note: A. Transient overexpression of FMRP-down-regulated global protein synthesis in cells. B. Transient interference of FMRP with siRNA-up-regulated global protein synthesis in cells. C. Transient overexpression of AAK1-up-regulated global protein synthesis in cells. D. Transient interference of AAK1 with siRNA-down-regulated global protein synthesis in cells.
Fig 4
Verification of AAK1 regulation on protein synthesis in cells by puromycin incorporation assays
WU Hao and HU Ronggui instructed the research. JIANG Guixian performed the experiments and data analyzing. The manuscript was drafted and revised by JIANG Guixian. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
All authors disclose no relevant conflict of interests.
CONNER S D, SCHMID S L. Identification of an adaptor-associated kinase, AAK1, as a regulator of clathrin-mediated endocytosis[J]. J Cell Biol, 2002, 156(5): 921-929.
AGAJANIAN M J, WALKER M P, AXTMAN A D, et al. WNT activates the AAK1 kinase to promote clathrin-mediated endocytosis of LRP6 and establish a negative feedback loop[J]. Cell Rep, 2019, 26(1): 79-93.e8.
GUPTA-ROSSI N, ORTICA S, MEAS-YEDID V, et al. The adaptor-associated kinase 1, AAK1, is a positive regulator of the Notch pathway[J]. J Biol Chem, 2011, 286(21): 18720-18730.
NEVEU G, ZIV-AV A, BAROUCH-BENTOV R, et al. AP2-associated protein kinase 1 and cyclin G-associated kinase regulate hepatitis C virus entry and are potential drug targets[J]. J Virol, 2015, 89(8): 4387-4404.
WANG C, WANG J, SHUAI L, et al. The serine/threonine kinase AP2-associated kinase 1 plays an important role in rabies virus entry[J]. Viruses, 2019, 12(1): E45.
MARTINEZ-GUALDA B, SCHOLS D, DE JONGHE S. A patent review of adaptor associated kinase 1 (AAK1) inhibitors (2013-present)[J]. Expert Opin Ther Pat, 2021, 31(10): 911-936.
KIRKPATRICK L L, MCILWAIN K A, NELSON D L. Comparative genomic sequence analysis of the FXR gene family: FMR1, FXR1, and FXR2[J]. Genomics, 2001, 78(3): 169-177.
VERKERK A J, PIERETTI M, SUTCLIFFE J S, et al. Identification of a gene (FMR-1) containing a CGG repeat coincident with a breakpoint cluster region exhibiting length variation in fragile X syndrome[J]. Cell, 1991, 65(5): 905-914.
SHAH S, MOLINARO G, LIU B, et al. FMRP control of ribosome translocation promotes chromatin modifications and alternative splicing of neuronal genes linked to autism[J]. Cell Rep, 2020, 30(13): 4459-4472.e6.
KORB E, HERRE M, ZUCKER-SCHARFF I, et al. Excess translation of epigenetic regulators contributes to fragile X syndrome and is alleviated by Brd4 inhibition[J]. Cell, 2017, 170(6): 1209-1223.e20.
DARNELL J C, VAN DRIESCHE S J, ZHANG C, et al. FMRP stalls ribosomal translocation on mRNAs linked to synaptic function and autism[J]. Cell, 2011, 146(2): 247-261.
RICHTER J D, BASSELL G J, KLANN E. Dysregulation and restoration of translational homeostasis in fragile X syndrome[J]. Nat Rev Neurosci, 2015, 16(10): 595-605.
EDENS B M, VISSERS C, SU J, et al. FMRP modulates neural differentiation through m6A-dependent mRNA nuclear export[J]. Cell Rep, 2019, 28(4): 845-854.e5.
TANG B, WANG T, WAN H, et al. Fmr1 deficiency promotes age-dependent alterations in the cortical synaptic proteome[J]. Proc Natl Acad Sci USA, 2015, 112(34): E4697-E4706.
LIAN J, ZHU X, DU J, et al. Extracellular vesicle-transmitted miR-671-5p alleviates lung inflammation and injury by regulating the AAK1/NF-κB axis[J].Mol Ther, 2023, 31(5): 1365-1382.
SORRELL F J, SZKLARZ M, ABDUL AZEEZ K R, et al. Family-wide structural analysis of human numb-associated protein kinases[J]. Structure, 2016, 24(3): 401-411.
HARTZ R A, AHUJA V T, NARA S J, et al. Discovery, structure-activity relationships, and in vivo evaluation of novel aryl amides as brain penetrant adaptor protein 2-associated kinase 1 (AAK1) inhibitors for the treatment of neuropathic pain[J]. J Med Chem, 2021, 64(15): 11090-11128.
XIN X, WANG Y, ZHANG L, et al. Development and therapeutic potential of adaptor-associated kinase 1 inhibitors in human multifaceted diseases[J]. Eur J Med Chem, 2023, 248: 115102.
ASAMITSU S, YABUKI Y, IKENOSHITA S, et al. CGG repeat RNA G-quadruplexes interact with FMRpolyG to cause neuronal dysfunction in fragile X-related tremor/ataxia syndrome[J]. Sci Adv, 2021, 7(3): eabd9440.
HALE C R, SAWICKA K, MORA K, et al. FMRP regulates mRNAs encoding distinct functions in the cell body and dendrites of CA1 pyramidal neurons[J]. Elife, 2021, 10: e71892.
MAURIN T, LEBRIGAND K, CASTAGNOLA S, et al. HITS-CLIP in various brain areas reveals new targets and new modalities of RNA binding by fragile X mental retardation protein[J]. Nucleic Acids Res, 2018, 46(12): 6344-6355.
BUDDIKA K, XU J, ARIYAPALA I S, et al. I-KCKT allows dissection-free RNA profiling of adult Drosophila intestinal progenitor cells[J]. Development, 2021, 148(1): dev196568.
MAZROUI R, HUOT M E, TREMBLAY S, et al. Trapping of messenger RNA by fragile X mental retardation protein into cytoplasmic granules induces translation repression[J]. Hum Mol Genet, 2002, 11(24): 3007-3017.
MONDAY H R, KHAROD S C, YOON Y J, et al. Presynaptic FMRP and local protein synthesis support structural and functional plasticity of glutamatergic axon terminals[J]. Neuron, 2022, 110(16): 2588-2606.e6.
KUAI L, ONG S E, MADISON J M, et al. AAK1 identified as an inhibitor of neuregulin-1/ErbB4-dependent neurotrophic factor signaling using integrative chemical genomics and proteomics[J]. Chem Biol, 2011, 18(7): 891-906.
... 衔接子相关蛋白激酶1(adaptor-associated protein kinase 1,AAK1)于2002年被首次鉴定,并发现其与网格蛋白介导的内吞作用(clathrin-mediated endocytosis,CME)中的关键分子衔接蛋白2(adaptor protein2,AP2)结合,通过磷酸化AP2的μ亚基(AP2 complex subunit μ,AP2M1)对CME过程进行调控[1-2].因此,AAK1也被称为AP2相关蛋白激酶1.目前,关于AAK1的生物学功能主要围绕CME展开.研究发现,在Wnt信号刺激下,AAK1通过加强AP2M1磷酸化程度,使低密度脂蛋白相关受体6(low-density lipoprotein receptor related protein 6,LRP6)通过CME途径从细胞膜转移到细胞质内[3];AAK1磷酸化内吞衔接蛋白Numb,使其从细胞膜通过CME途径进入内体[4].除了磷酸化作用外,AAK1还以非磷酸化方式直接结合并稳定活化形式的神经源性基因座缺口同源蛋白1(neurogenic locus notch homolog protein 1,Notch1),使其进入内体并增强其转录活性[5].AAK1也参与疾病的发生和发展.超氧化物歧化酶1蛋白(superoxide dismutase 1,SOD1)突变(SOD1G85R)与肌萎缩侧索硬化有关,而AAK1仅与SOD1G85R结合且不与SOD1WT结合,并且AAK1蛋白水平在肌萎缩侧索硬化患者中较正常人低[6].部分病毒通过CME途径进入人体细胞,而AAK1参与调节丙型肝炎病毒[7]、登革热病毒[8]、狂犬病病毒[9]以及在全球范围内流行的新型冠状病毒[10]的进入与细胞间的扩散.因此,也有针对AAK1开展的各类化合物的研发[11-12].鉴于AAK1对生理、病理过程均有广泛的调控作用,有必要对AAK1开展更深入的研究,探索其更多的分子机制. ...
... 衔接子相关蛋白激酶1(adaptor-associated protein kinase 1,AAK1)于2002年被首次鉴定,并发现其与网格蛋白介导的内吞作用(clathrin-mediated endocytosis,CME)中的关键分子衔接蛋白2(adaptor protein2,AP2)结合,通过磷酸化AP2的μ亚基(AP2 complex subunit μ,AP2M1)对CME过程进行调控[1-2].因此,AAK1也被称为AP2相关蛋白激酶1.目前,关于AAK1的生物学功能主要围绕CME展开.研究发现,在Wnt信号刺激下,AAK1通过加强AP2M1磷酸化程度,使低密度脂蛋白相关受体6(low-density lipoprotein receptor related protein 6,LRP6)通过CME途径从细胞膜转移到细胞质内[3];AAK1磷酸化内吞衔接蛋白Numb,使其从细胞膜通过CME途径进入内体[4].除了磷酸化作用外,AAK1还以非磷酸化方式直接结合并稳定活化形式的神经源性基因座缺口同源蛋白1(neurogenic locus notch homolog protein 1,Notch1),使其进入内体并增强其转录活性[5].AAK1也参与疾病的发生和发展.超氧化物歧化酶1蛋白(superoxide dismutase 1,SOD1)突变(SOD1G85R)与肌萎缩侧索硬化有关,而AAK1仅与SOD1G85R结合且不与SOD1WT结合,并且AAK1蛋白水平在肌萎缩侧索硬化患者中较正常人低[6].部分病毒通过CME途径进入人体细胞,而AAK1参与调节丙型肝炎病毒[7]、登革热病毒[8]、狂犬病病毒[9]以及在全球范围内流行的新型冠状病毒[10]的进入与细胞间的扩散.因此,也有针对AAK1开展的各类化合物的研发[11-12].鉴于AAK1对生理、病理过程均有广泛的调控作用,有必要对AAK1开展更深入的研究,探索其更多的分子机制. ...
1
... 衔接子相关蛋白激酶1(adaptor-associated protein kinase 1,AAK1)于2002年被首次鉴定,并发现其与网格蛋白介导的内吞作用(clathrin-mediated endocytosis,CME)中的关键分子衔接蛋白2(adaptor protein2,AP2)结合,通过磷酸化AP2的μ亚基(AP2 complex subunit μ,AP2M1)对CME过程进行调控[1-2].因此,AAK1也被称为AP2相关蛋白激酶1.目前,关于AAK1的生物学功能主要围绕CME展开.研究发现,在Wnt信号刺激下,AAK1通过加强AP2M1磷酸化程度,使低密度脂蛋白相关受体6(low-density lipoprotein receptor related protein 6,LRP6)通过CME途径从细胞膜转移到细胞质内[3];AAK1磷酸化内吞衔接蛋白Numb,使其从细胞膜通过CME途径进入内体[4].除了磷酸化作用外,AAK1还以非磷酸化方式直接结合并稳定活化形式的神经源性基因座缺口同源蛋白1(neurogenic locus notch homolog protein 1,Notch1),使其进入内体并增强其转录活性[5].AAK1也参与疾病的发生和发展.超氧化物歧化酶1蛋白(superoxide dismutase 1,SOD1)突变(SOD1G85R)与肌萎缩侧索硬化有关,而AAK1仅与SOD1G85R结合且不与SOD1WT结合,并且AAK1蛋白水平在肌萎缩侧索硬化患者中较正常人低[6].部分病毒通过CME途径进入人体细胞,而AAK1参与调节丙型肝炎病毒[7]、登革热病毒[8]、狂犬病病毒[9]以及在全球范围内流行的新型冠状病毒[10]的进入与细胞间的扩散.因此,也有针对AAK1开展的各类化合物的研发[11-12].鉴于AAK1对生理、病理过程均有广泛的调控作用,有必要对AAK1开展更深入的研究,探索其更多的分子机制. ...
1
... 衔接子相关蛋白激酶1(adaptor-associated protein kinase 1,AAK1)于2002年被首次鉴定,并发现其与网格蛋白介导的内吞作用(clathrin-mediated endocytosis,CME)中的关键分子衔接蛋白2(adaptor protein2,AP2)结合,通过磷酸化AP2的μ亚基(AP2 complex subunit μ,AP2M1)对CME过程进行调控[1-2].因此,AAK1也被称为AP2相关蛋白激酶1.目前,关于AAK1的生物学功能主要围绕CME展开.研究发现,在Wnt信号刺激下,AAK1通过加强AP2M1磷酸化程度,使低密度脂蛋白相关受体6(low-density lipoprotein receptor related protein 6,LRP6)通过CME途径从细胞膜转移到细胞质内[3];AAK1磷酸化内吞衔接蛋白Numb,使其从细胞膜通过CME途径进入内体[4].除了磷酸化作用外,AAK1还以非磷酸化方式直接结合并稳定活化形式的神经源性基因座缺口同源蛋白1(neurogenic locus notch homolog protein 1,Notch1),使其进入内体并增强其转录活性[5].AAK1也参与疾病的发生和发展.超氧化物歧化酶1蛋白(superoxide dismutase 1,SOD1)突变(SOD1G85R)与肌萎缩侧索硬化有关,而AAK1仅与SOD1G85R结合且不与SOD1WT结合,并且AAK1蛋白水平在肌萎缩侧索硬化患者中较正常人低[6].部分病毒通过CME途径进入人体细胞,而AAK1参与调节丙型肝炎病毒[7]、登革热病毒[8]、狂犬病病毒[9]以及在全球范围内流行的新型冠状病毒[10]的进入与细胞间的扩散.因此,也有针对AAK1开展的各类化合物的研发[11-12].鉴于AAK1对生理、病理过程均有广泛的调控作用,有必要对AAK1开展更深入的研究,探索其更多的分子机制. ...
1
... 衔接子相关蛋白激酶1(adaptor-associated protein kinase 1,AAK1)于2002年被首次鉴定,并发现其与网格蛋白介导的内吞作用(clathrin-mediated endocytosis,CME)中的关键分子衔接蛋白2(adaptor protein2,AP2)结合,通过磷酸化AP2的μ亚基(AP2 complex subunit μ,AP2M1)对CME过程进行调控[1-2].因此,AAK1也被称为AP2相关蛋白激酶1.目前,关于AAK1的生物学功能主要围绕CME展开.研究发现,在Wnt信号刺激下,AAK1通过加强AP2M1磷酸化程度,使低密度脂蛋白相关受体6(low-density lipoprotein receptor related protein 6,LRP6)通过CME途径从细胞膜转移到细胞质内[3];AAK1磷酸化内吞衔接蛋白Numb,使其从细胞膜通过CME途径进入内体[4].除了磷酸化作用外,AAK1还以非磷酸化方式直接结合并稳定活化形式的神经源性基因座缺口同源蛋白1(neurogenic locus notch homolog protein 1,Notch1),使其进入内体并增强其转录活性[5].AAK1也参与疾病的发生和发展.超氧化物歧化酶1蛋白(superoxide dismutase 1,SOD1)突变(SOD1G85R)与肌萎缩侧索硬化有关,而AAK1仅与SOD1G85R结合且不与SOD1WT结合,并且AAK1蛋白水平在肌萎缩侧索硬化患者中较正常人低[6].部分病毒通过CME途径进入人体细胞,而AAK1参与调节丙型肝炎病毒[7]、登革热病毒[8]、狂犬病病毒[9]以及在全球范围内流行的新型冠状病毒[10]的进入与细胞间的扩散.因此,也有针对AAK1开展的各类化合物的研发[11-12].鉴于AAK1对生理、病理过程均有广泛的调控作用,有必要对AAK1开展更深入的研究,探索其更多的分子机制. ...
1
... 衔接子相关蛋白激酶1(adaptor-associated protein kinase 1,AAK1)于2002年被首次鉴定,并发现其与网格蛋白介导的内吞作用(clathrin-mediated endocytosis,CME)中的关键分子衔接蛋白2(adaptor protein2,AP2)结合,通过磷酸化AP2的μ亚基(AP2 complex subunit μ,AP2M1)对CME过程进行调控[1-2].因此,AAK1也被称为AP2相关蛋白激酶1.目前,关于AAK1的生物学功能主要围绕CME展开.研究发现,在Wnt信号刺激下,AAK1通过加强AP2M1磷酸化程度,使低密度脂蛋白相关受体6(low-density lipoprotein receptor related protein 6,LRP6)通过CME途径从细胞膜转移到细胞质内[3];AAK1磷酸化内吞衔接蛋白Numb,使其从细胞膜通过CME途径进入内体[4].除了磷酸化作用外,AAK1还以非磷酸化方式直接结合并稳定活化形式的神经源性基因座缺口同源蛋白1(neurogenic locus notch homolog protein 1,Notch1),使其进入内体并增强其转录活性[5].AAK1也参与疾病的发生和发展.超氧化物歧化酶1蛋白(superoxide dismutase 1,SOD1)突变(SOD1G85R)与肌萎缩侧索硬化有关,而AAK1仅与SOD1G85R结合且不与SOD1WT结合,并且AAK1蛋白水平在肌萎缩侧索硬化患者中较正常人低[6].部分病毒通过CME途径进入人体细胞,而AAK1参与调节丙型肝炎病毒[7]、登革热病毒[8]、狂犬病病毒[9]以及在全球范围内流行的新型冠状病毒[10]的进入与细胞间的扩散.因此,也有针对AAK1开展的各类化合物的研发[11-12].鉴于AAK1对生理、病理过程均有广泛的调控作用,有必要对AAK1开展更深入的研究,探索其更多的分子机制. ...
1
... 衔接子相关蛋白激酶1(adaptor-associated protein kinase 1,AAK1)于2002年被首次鉴定,并发现其与网格蛋白介导的内吞作用(clathrin-mediated endocytosis,CME)中的关键分子衔接蛋白2(adaptor protein2,AP2)结合,通过磷酸化AP2的μ亚基(AP2 complex subunit μ,AP2M1)对CME过程进行调控[1-2].因此,AAK1也被称为AP2相关蛋白激酶1.目前,关于AAK1的生物学功能主要围绕CME展开.研究发现,在Wnt信号刺激下,AAK1通过加强AP2M1磷酸化程度,使低密度脂蛋白相关受体6(low-density lipoprotein receptor related protein 6,LRP6)通过CME途径从细胞膜转移到细胞质内[3];AAK1磷酸化内吞衔接蛋白Numb,使其从细胞膜通过CME途径进入内体[4].除了磷酸化作用外,AAK1还以非磷酸化方式直接结合并稳定活化形式的神经源性基因座缺口同源蛋白1(neurogenic locus notch homolog protein 1,Notch1),使其进入内体并增强其转录活性[5].AAK1也参与疾病的发生和发展.超氧化物歧化酶1蛋白(superoxide dismutase 1,SOD1)突变(SOD1G85R)与肌萎缩侧索硬化有关,而AAK1仅与SOD1G85R结合且不与SOD1WT结合,并且AAK1蛋白水平在肌萎缩侧索硬化患者中较正常人低[6].部分病毒通过CME途径进入人体细胞,而AAK1参与调节丙型肝炎病毒[7]、登革热病毒[8]、狂犬病病毒[9]以及在全球范围内流行的新型冠状病毒[10]的进入与细胞间的扩散.因此,也有针对AAK1开展的各类化合物的研发[11-12].鉴于AAK1对生理、病理过程均有广泛的调控作用,有必要对AAK1开展更深入的研究,探索其更多的分子机制. ...
1
... 衔接子相关蛋白激酶1(adaptor-associated protein kinase 1,AAK1)于2002年被首次鉴定,并发现其与网格蛋白介导的内吞作用(clathrin-mediated endocytosis,CME)中的关键分子衔接蛋白2(adaptor protein2,AP2)结合,通过磷酸化AP2的μ亚基(AP2 complex subunit μ,AP2M1)对CME过程进行调控[1-2].因此,AAK1也被称为AP2相关蛋白激酶1.目前,关于AAK1的生物学功能主要围绕CME展开.研究发现,在Wnt信号刺激下,AAK1通过加强AP2M1磷酸化程度,使低密度脂蛋白相关受体6(low-density lipoprotein receptor related protein 6,LRP6)通过CME途径从细胞膜转移到细胞质内[3];AAK1磷酸化内吞衔接蛋白Numb,使其从细胞膜通过CME途径进入内体[4].除了磷酸化作用外,AAK1还以非磷酸化方式直接结合并稳定活化形式的神经源性基因座缺口同源蛋白1(neurogenic locus notch homolog protein 1,Notch1),使其进入内体并增强其转录活性[5].AAK1也参与疾病的发生和发展.超氧化物歧化酶1蛋白(superoxide dismutase 1,SOD1)突变(SOD1G85R)与肌萎缩侧索硬化有关,而AAK1仅与SOD1G85R结合且不与SOD1WT结合,并且AAK1蛋白水平在肌萎缩侧索硬化患者中较正常人低[6].部分病毒通过CME途径进入人体细胞,而AAK1参与调节丙型肝炎病毒[7]、登革热病毒[8]、狂犬病病毒[9]以及在全球范围内流行的新型冠状病毒[10]的进入与细胞间的扩散.因此,也有针对AAK1开展的各类化合物的研发[11-12].鉴于AAK1对生理、病理过程均有广泛的调控作用,有必要对AAK1开展更深入的研究,探索其更多的分子机制. ...
1
... 衔接子相关蛋白激酶1(adaptor-associated protein kinase 1,AAK1)于2002年被首次鉴定,并发现其与网格蛋白介导的内吞作用(clathrin-mediated endocytosis,CME)中的关键分子衔接蛋白2(adaptor protein2,AP2)结合,通过磷酸化AP2的μ亚基(AP2 complex subunit μ,AP2M1)对CME过程进行调控[1-2].因此,AAK1也被称为AP2相关蛋白激酶1.目前,关于AAK1的生物学功能主要围绕CME展开.研究发现,在Wnt信号刺激下,AAK1通过加强AP2M1磷酸化程度,使低密度脂蛋白相关受体6(low-density lipoprotein receptor related protein 6,LRP6)通过CME途径从细胞膜转移到细胞质内[3];AAK1磷酸化内吞衔接蛋白Numb,使其从细胞膜通过CME途径进入内体[4].除了磷酸化作用外,AAK1还以非磷酸化方式直接结合并稳定活化形式的神经源性基因座缺口同源蛋白1(neurogenic locus notch homolog protein 1,Notch1),使其进入内体并增强其转录活性[5].AAK1也参与疾病的发生和发展.超氧化物歧化酶1蛋白(superoxide dismutase 1,SOD1)突变(SOD1G85R)与肌萎缩侧索硬化有关,而AAK1仅与SOD1G85R结合且不与SOD1WT结合,并且AAK1蛋白水平在肌萎缩侧索硬化患者中较正常人低[6].部分病毒通过CME途径进入人体细胞,而AAK1参与调节丙型肝炎病毒[7]、登革热病毒[8]、狂犬病病毒[9]以及在全球范围内流行的新型冠状病毒[10]的进入与细胞间的扩散.因此,也有针对AAK1开展的各类化合物的研发[11-12].鉴于AAK1对生理、病理过程均有广泛的调控作用,有必要对AAK1开展更深入的研究,探索其更多的分子机制. ...
1
... 衔接子相关蛋白激酶1(adaptor-associated protein kinase 1,AAK1)于2002年被首次鉴定,并发现其与网格蛋白介导的内吞作用(clathrin-mediated endocytosis,CME)中的关键分子衔接蛋白2(adaptor protein2,AP2)结合,通过磷酸化AP2的μ亚基(AP2 complex subunit μ,AP2M1)对CME过程进行调控[1-2].因此,AAK1也被称为AP2相关蛋白激酶1.目前,关于AAK1的生物学功能主要围绕CME展开.研究发现,在Wnt信号刺激下,AAK1通过加强AP2M1磷酸化程度,使低密度脂蛋白相关受体6(low-density lipoprotein receptor related protein 6,LRP6)通过CME途径从细胞膜转移到细胞质内[3];AAK1磷酸化内吞衔接蛋白Numb,使其从细胞膜通过CME途径进入内体[4].除了磷酸化作用外,AAK1还以非磷酸化方式直接结合并稳定活化形式的神经源性基因座缺口同源蛋白1(neurogenic locus notch homolog protein 1,Notch1),使其进入内体并增强其转录活性[5].AAK1也参与疾病的发生和发展.超氧化物歧化酶1蛋白(superoxide dismutase 1,SOD1)突变(SOD1G85R)与肌萎缩侧索硬化有关,而AAK1仅与SOD1G85R结合且不与SOD1WT结合,并且AAK1蛋白水平在肌萎缩侧索硬化患者中较正常人低[6].部分病毒通过CME途径进入人体细胞,而AAK1参与调节丙型肝炎病毒[7]、登革热病毒[8]、狂犬病病毒[9]以及在全球范围内流行的新型冠状病毒[10]的进入与细胞间的扩散.因此,也有针对AAK1开展的各类化合物的研发[11-12].鉴于AAK1对生理、病理过程均有广泛的调控作用,有必要对AAK1开展更深入的研究,探索其更多的分子机制. ...
1
... 衔接子相关蛋白激酶1(adaptor-associated protein kinase 1,AAK1)于2002年被首次鉴定,并发现其与网格蛋白介导的内吞作用(clathrin-mediated endocytosis,CME)中的关键分子衔接蛋白2(adaptor protein2,AP2)结合,通过磷酸化AP2的μ亚基(AP2 complex subunit μ,AP2M1)对CME过程进行调控[1-2].因此,AAK1也被称为AP2相关蛋白激酶1.目前,关于AAK1的生物学功能主要围绕CME展开.研究发现,在Wnt信号刺激下,AAK1通过加强AP2M1磷酸化程度,使低密度脂蛋白相关受体6(low-density lipoprotein receptor related protein 6,LRP6)通过CME途径从细胞膜转移到细胞质内[3];AAK1磷酸化内吞衔接蛋白Numb,使其从细胞膜通过CME途径进入内体[4].除了磷酸化作用外,AAK1还以非磷酸化方式直接结合并稳定活化形式的神经源性基因座缺口同源蛋白1(neurogenic locus notch homolog protein 1,Notch1),使其进入内体并增强其转录活性[5].AAK1也参与疾病的发生和发展.超氧化物歧化酶1蛋白(superoxide dismutase 1,SOD1)突变(SOD1G85R)与肌萎缩侧索硬化有关,而AAK1仅与SOD1G85R结合且不与SOD1WT结合,并且AAK1蛋白水平在肌萎缩侧索硬化患者中较正常人低[6].部分病毒通过CME途径进入人体细胞,而AAK1参与调节丙型肝炎病毒[7]、登革热病毒[8]、狂犬病病毒[9]以及在全球范围内流行的新型冠状病毒[10]的进入与细胞间的扩散.因此,也有针对AAK1开展的各类化合物的研发[11-12].鉴于AAK1对生理、病理过程均有广泛的调控作用,有必要对AAK1开展更深入的研究,探索其更多的分子机制. ...
1
... 衔接子相关蛋白激酶1(adaptor-associated protein kinase 1,AAK1)于2002年被首次鉴定,并发现其与网格蛋白介导的内吞作用(clathrin-mediated endocytosis,CME)中的关键分子衔接蛋白2(adaptor protein2,AP2)结合,通过磷酸化AP2的μ亚基(AP2 complex subunit μ,AP2M1)对CME过程进行调控[1-2].因此,AAK1也被称为AP2相关蛋白激酶1.目前,关于AAK1的生物学功能主要围绕CME展开.研究发现,在Wnt信号刺激下,AAK1通过加强AP2M1磷酸化程度,使低密度脂蛋白相关受体6(low-density lipoprotein receptor related protein 6,LRP6)通过CME途径从细胞膜转移到细胞质内[3];AAK1磷酸化内吞衔接蛋白Numb,使其从细胞膜通过CME途径进入内体[4].除了磷酸化作用外,AAK1还以非磷酸化方式直接结合并稳定活化形式的神经源性基因座缺口同源蛋白1(neurogenic locus notch homolog protein 1,Notch1),使其进入内体并增强其转录活性[5].AAK1也参与疾病的发生和发展.超氧化物歧化酶1蛋白(superoxide dismutase 1,SOD1)突变(SOD1G85R)与肌萎缩侧索硬化有关,而AAK1仅与SOD1G85R结合且不与SOD1WT结合,并且AAK1蛋白水平在肌萎缩侧索硬化患者中较正常人低[6].部分病毒通过CME途径进入人体细胞,而AAK1参与调节丙型肝炎病毒[7]、登革热病毒[8]、狂犬病病毒[9]以及在全球范围内流行的新型冠状病毒[10]的进入与细胞间的扩散.因此,也有针对AAK1开展的各类化合物的研发[11-12].鉴于AAK1对生理、病理过程均有广泛的调控作用,有必要对AAK1开展更深入的研究,探索其更多的分子机制. ...
1
... 脆性X智力低下蛋白(fragile X mental retardation protein 1,FMRP)是脆性X相关(fragile X mental retardation syndrome-related,FXR)蛋白家族的一员,其与脆性X相关蛋白1(FXR protein 1,FXR1)和FXR2具有显著的同源性,在结构、功能上相似[13].FXR蛋白家族的表达异常是脆性X综合征的直接病因.该病是以智力低下和自闭症样行为为主的一类遗传性疾病,其中以FMRP异常为主[14].FMRP的功能繁多,可以从多个层面对机体进行调控:① 在DNA层面,调控转录因子与染色质修饰酶的表达与活性[15-16].② 在蛋白层面,通过直接与离子通道蛋白结合进而影响其兴奋性[17].③ 在RNA层面,由于FMRP是RNA结合蛋白,FMRP还通过诱导核糖体停滞[18]、与RNA直接结合调控其稳定性[19]、调控RNA在细胞内定位[20]等方式参与调控RNA代谢.运用定量质谱比较青春期和成年期的Fmrp敲除和野生型2种小鼠皮层突触的蛋白质表达情况发现,Fmrp敲除使青春期小鼠大量突触蛋白上调,而在成年小鼠中Fmrp敲除影响较小,表明FMRP的调控还具有时间特异性[21].由此可见,FMRP的分子机制是十分复杂的.目前,对FMRP的认识相较于FXR1、FXR2更加深入,研究上常以对FMRP的研究为基础反推至FXR1与FXR2. ...
1
... 脆性X智力低下蛋白(fragile X mental retardation protein 1,FMRP)是脆性X相关(fragile X mental retardation syndrome-related,FXR)蛋白家族的一员,其与脆性X相关蛋白1(FXR protein 1,FXR1)和FXR2具有显著的同源性,在结构、功能上相似[13].FXR蛋白家族的表达异常是脆性X综合征的直接病因.该病是以智力低下和自闭症样行为为主的一类遗传性疾病,其中以FMRP异常为主[14].FMRP的功能繁多,可以从多个层面对机体进行调控:① 在DNA层面,调控转录因子与染色质修饰酶的表达与活性[15-16].② 在蛋白层面,通过直接与离子通道蛋白结合进而影响其兴奋性[17].③ 在RNA层面,由于FMRP是RNA结合蛋白,FMRP还通过诱导核糖体停滞[18]、与RNA直接结合调控其稳定性[19]、调控RNA在细胞内定位[20]等方式参与调控RNA代谢.运用定量质谱比较青春期和成年期的Fmrp敲除和野生型2种小鼠皮层突触的蛋白质表达情况发现,Fmrp敲除使青春期小鼠大量突触蛋白上调,而在成年小鼠中Fmrp敲除影响较小,表明FMRP的调控还具有时间特异性[21].由此可见,FMRP的分子机制是十分复杂的.目前,对FMRP的认识相较于FXR1、FXR2更加深入,研究上常以对FMRP的研究为基础反推至FXR1与FXR2. ...
1
... 脆性X智力低下蛋白(fragile X mental retardation protein 1,FMRP)是脆性X相关(fragile X mental retardation syndrome-related,FXR)蛋白家族的一员,其与脆性X相关蛋白1(FXR protein 1,FXR1)和FXR2具有显著的同源性,在结构、功能上相似[13].FXR蛋白家族的表达异常是脆性X综合征的直接病因.该病是以智力低下和自闭症样行为为主的一类遗传性疾病,其中以FMRP异常为主[14].FMRP的功能繁多,可以从多个层面对机体进行调控:① 在DNA层面,调控转录因子与染色质修饰酶的表达与活性[15-16].② 在蛋白层面,通过直接与离子通道蛋白结合进而影响其兴奋性[17].③ 在RNA层面,由于FMRP是RNA结合蛋白,FMRP还通过诱导核糖体停滞[18]、与RNA直接结合调控其稳定性[19]、调控RNA在细胞内定位[20]等方式参与调控RNA代谢.运用定量质谱比较青春期和成年期的Fmrp敲除和野生型2种小鼠皮层突触的蛋白质表达情况发现,Fmrp敲除使青春期小鼠大量突触蛋白上调,而在成年小鼠中Fmrp敲除影响较小,表明FMRP的调控还具有时间特异性[21].由此可见,FMRP的分子机制是十分复杂的.目前,对FMRP的认识相较于FXR1、FXR2更加深入,研究上常以对FMRP的研究为基础反推至FXR1与FXR2. ...
1
... 脆性X智力低下蛋白(fragile X mental retardation protein 1,FMRP)是脆性X相关(fragile X mental retardation syndrome-related,FXR)蛋白家族的一员,其与脆性X相关蛋白1(FXR protein 1,FXR1)和FXR2具有显著的同源性,在结构、功能上相似[13].FXR蛋白家族的表达异常是脆性X综合征的直接病因.该病是以智力低下和自闭症样行为为主的一类遗传性疾病,其中以FMRP异常为主[14].FMRP的功能繁多,可以从多个层面对机体进行调控:① 在DNA层面,调控转录因子与染色质修饰酶的表达与活性[15-16].② 在蛋白层面,通过直接与离子通道蛋白结合进而影响其兴奋性[17].③ 在RNA层面,由于FMRP是RNA结合蛋白,FMRP还通过诱导核糖体停滞[18]、与RNA直接结合调控其稳定性[19]、调控RNA在细胞内定位[20]等方式参与调控RNA代谢.运用定量质谱比较青春期和成年期的Fmrp敲除和野生型2种小鼠皮层突触的蛋白质表达情况发现,Fmrp敲除使青春期小鼠大量突触蛋白上调,而在成年小鼠中Fmrp敲除影响较小,表明FMRP的调控还具有时间特异性[21].由此可见,FMRP的分子机制是十分复杂的.目前,对FMRP的认识相较于FXR1、FXR2更加深入,研究上常以对FMRP的研究为基础反推至FXR1与FXR2. ...
1
... 脆性X智力低下蛋白(fragile X mental retardation protein 1,FMRP)是脆性X相关(fragile X mental retardation syndrome-related,FXR)蛋白家族的一员,其与脆性X相关蛋白1(FXR protein 1,FXR1)和FXR2具有显著的同源性,在结构、功能上相似[13].FXR蛋白家族的表达异常是脆性X综合征的直接病因.该病是以智力低下和自闭症样行为为主的一类遗传性疾病,其中以FMRP异常为主[14].FMRP的功能繁多,可以从多个层面对机体进行调控:① 在DNA层面,调控转录因子与染色质修饰酶的表达与活性[15-16].② 在蛋白层面,通过直接与离子通道蛋白结合进而影响其兴奋性[17].③ 在RNA层面,由于FMRP是RNA结合蛋白,FMRP还通过诱导核糖体停滞[18]、与RNA直接结合调控其稳定性[19]、调控RNA在细胞内定位[20]等方式参与调控RNA代谢.运用定量质谱比较青春期和成年期的Fmrp敲除和野生型2种小鼠皮层突触的蛋白质表达情况发现,Fmrp敲除使青春期小鼠大量突触蛋白上调,而在成年小鼠中Fmrp敲除影响较小,表明FMRP的调控还具有时间特异性[21].由此可见,FMRP的分子机制是十分复杂的.目前,对FMRP的认识相较于FXR1、FXR2更加深入,研究上常以对FMRP的研究为基础反推至FXR1与FXR2. ...
2
... 脆性X智力低下蛋白(fragile X mental retardation protein 1,FMRP)是脆性X相关(fragile X mental retardation syndrome-related,FXR)蛋白家族的一员,其与脆性X相关蛋白1(FXR protein 1,FXR1)和FXR2具有显著的同源性,在结构、功能上相似[13].FXR蛋白家族的表达异常是脆性X综合征的直接病因.该病是以智力低下和自闭症样行为为主的一类遗传性疾病,其中以FMRP异常为主[14].FMRP的功能繁多,可以从多个层面对机体进行调控:① 在DNA层面,调控转录因子与染色质修饰酶的表达与活性[15-16].② 在蛋白层面,通过直接与离子通道蛋白结合进而影响其兴奋性[17].③ 在RNA层面,由于FMRP是RNA结合蛋白,FMRP还通过诱导核糖体停滞[18]、与RNA直接结合调控其稳定性[19]、调控RNA在细胞内定位[20]等方式参与调控RNA代谢.运用定量质谱比较青春期和成年期的Fmrp敲除和野生型2种小鼠皮层突触的蛋白质表达情况发现,Fmrp敲除使青春期小鼠大量突触蛋白上调,而在成年小鼠中Fmrp敲除影响较小,表明FMRP的调控还具有时间特异性[21].由此可见,FMRP的分子机制是十分复杂的.目前,对FMRP的认识相较于FXR1、FXR2更加深入,研究上常以对FMRP的研究为基础反推至FXR1与FXR2. ...
... 脆性X智力低下蛋白(fragile X mental retardation protein 1,FMRP)是脆性X相关(fragile X mental retardation syndrome-related,FXR)蛋白家族的一员,其与脆性X相关蛋白1(FXR protein 1,FXR1)和FXR2具有显著的同源性,在结构、功能上相似[13].FXR蛋白家族的表达异常是脆性X综合征的直接病因.该病是以智力低下和自闭症样行为为主的一类遗传性疾病,其中以FMRP异常为主[14].FMRP的功能繁多,可以从多个层面对机体进行调控:① 在DNA层面,调控转录因子与染色质修饰酶的表达与活性[15-16].② 在蛋白层面,通过直接与离子通道蛋白结合进而影响其兴奋性[17].③ 在RNA层面,由于FMRP是RNA结合蛋白,FMRP还通过诱导核糖体停滞[18]、与RNA直接结合调控其稳定性[19]、调控RNA在细胞内定位[20]等方式参与调控RNA代谢.运用定量质谱比较青春期和成年期的Fmrp敲除和野生型2种小鼠皮层突触的蛋白质表达情况发现,Fmrp敲除使青春期小鼠大量突触蛋白上调,而在成年小鼠中Fmrp敲除影响较小,表明FMRP的调控还具有时间特异性[21].由此可见,FMRP的分子机制是十分复杂的.目前,对FMRP的认识相较于FXR1、FXR2更加深入,研究上常以对FMRP的研究为基础反推至FXR1与FXR2. ...
1
... 脆性X智力低下蛋白(fragile X mental retardation protein 1,FMRP)是脆性X相关(fragile X mental retardation syndrome-related,FXR)蛋白家族的一员,其与脆性X相关蛋白1(FXR protein 1,FXR1)和FXR2具有显著的同源性,在结构、功能上相似[13].FXR蛋白家族的表达异常是脆性X综合征的直接病因.该病是以智力低下和自闭症样行为为主的一类遗传性疾病,其中以FMRP异常为主[14].FMRP的功能繁多,可以从多个层面对机体进行调控:① 在DNA层面,调控转录因子与染色质修饰酶的表达与活性[15-16].② 在蛋白层面,通过直接与离子通道蛋白结合进而影响其兴奋性[17].③ 在RNA层面,由于FMRP是RNA结合蛋白,FMRP还通过诱导核糖体停滞[18]、与RNA直接结合调控其稳定性[19]、调控RNA在细胞内定位[20]等方式参与调控RNA代谢.运用定量质谱比较青春期和成年期的Fmrp敲除和野生型2种小鼠皮层突触的蛋白质表达情况发现,Fmrp敲除使青春期小鼠大量突触蛋白上调,而在成年小鼠中Fmrp敲除影响较小,表明FMRP的调控还具有时间特异性[21].由此可见,FMRP的分子机制是十分复杂的.目前,对FMRP的认识相较于FXR1、FXR2更加深入,研究上常以对FMRP的研究为基础反推至FXR1与FXR2. ...
1
... 脆性X智力低下蛋白(fragile X mental retardation protein 1,FMRP)是脆性X相关(fragile X mental retardation syndrome-related,FXR)蛋白家族的一员,其与脆性X相关蛋白1(FXR protein 1,FXR1)和FXR2具有显著的同源性,在结构、功能上相似[13].FXR蛋白家族的表达异常是脆性X综合征的直接病因.该病是以智力低下和自闭症样行为为主的一类遗传性疾病,其中以FMRP异常为主[14].FMRP的功能繁多,可以从多个层面对机体进行调控:① 在DNA层面,调控转录因子与染色质修饰酶的表达与活性[15-16].② 在蛋白层面,通过直接与离子通道蛋白结合进而影响其兴奋性[17].③ 在RNA层面,由于FMRP是RNA结合蛋白,FMRP还通过诱导核糖体停滞[18]、与RNA直接结合调控其稳定性[19]、调控RNA在细胞内定位[20]等方式参与调控RNA代谢.运用定量质谱比较青春期和成年期的Fmrp敲除和野生型2种小鼠皮层突触的蛋白质表达情况发现,Fmrp敲除使青春期小鼠大量突触蛋白上调,而在成年小鼠中Fmrp敲除影响较小,表明FMRP的调控还具有时间特异性[21].由此可见,FMRP的分子机制是十分复杂的.目前,对FMRP的认识相较于FXR1、FXR2更加深入,研究上常以对FMRP的研究为基础反推至FXR1与FXR2. ...





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}