Structural analysis of full-length lysine acetyltransferase 7 by cryo-electron microscopy
ZHENG Guopei,1, CAO Qin2, SHEN Jianfeng,1
1.Department of Ophthalmology, Shanghai Ninth People's Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai 200011, China
2.Bio-X Institutes, Key Laboratory for the Genetics of Developmental and Neuropsychiatric Disorders, Ministry of Education, Shanghai Jiao Tong University, Shanghai 200030, China
Objective ·To analyze the full-length protein structure of human-derived lysine acetyltransferase 7 (KAT7) using cryo-electron microscopy (Cryo-EM) and to obtain the profile information of human-derived KAT7. Methods ·The recombinant protein expression plasmid pGEX-4T1-GST-KAT7 was constructed by using the pGEX-4T1 vector and the full-length gene of human-derived KAT7, and the KAT7 protein was expressed in the prokaryotic protein expression system BL21 (DE3). The GST-KAT7 recombinant protein was obtained by using GST affinity chromatography. After removing the GST protein tag with TEV protease, KAT7 was further isolated and purified by HiLoad 16/600 Superdex 75 pg volume exclusion chromatography. The obtained protein samples were identified by Western blotting, and the samples were screened. The protein morphology was observed under negative-stain electron microscopy, and data were collected by using Cryo-EM. The protein particles were selected and the spatial structure of the full-length KAT7 was analyzed with the Cryo-EM analysis software CryoSparc. The MYST structural domain model (5GK9) in the Protein Data Bank (PDB) and AlphaFold prediction model of KAT7 were matched with the generated structural model by UCSF Chimera software. Results ·The full-length protein of human-derived KAT7 was successfully purified by affinity chromatography, and high purity KAT7 was obtained by volume exclusion chromatography. After identifying KAT7 by Western blotting, the spatial structure of KAT7 full-length protein was initially resolved by Cryo-EM and single-particle reconstruction techniques, and a preliminary three-dimensional structure model with a resolution of about 10 Å was obtained by three-dimensional optimization. The spatial structure of KAT7 full-length protein was irregular and semi-loop-shaped, and the existing MYST domain model (PDB: 5GK9) can be matched into the C-terminal part of the KAT7 full-length model. The adjusted AlphaFold prediction model can also match the KAT7 full-length structure model. Conclusion ·A preliminary analysis of the spatial structure model of full-length protein of human-derived KAT7 is performed by using Cryo-EM.
人源性KAT7具有611个氨基酸残基,主要由2个结构域组成:N端结构域包含1个短的锌指DNA结合域,它主要与MCM2(minichromosome maintenance complex component 2)和ORC1(origin recognition complex subunit 1)相互作用;C端为MYST乙酰基转移酶结构域,主要负责结合乙酰辅酶A并催化对应的乙酰化反应[16]。MYST结构域是MYST家族共有的乙酰转移酶结构域,其在多种HAT蛋白如MYST1(MOF/KAT8)、MYST2(HBO1/KAT7)和MYST3(MoZ/KAT6A)中高度保守[17]。HAT通常与支架蛋白和其他辅助蛋白相互作用,形成HAT复合物并发挥功能[18]。已经鉴定的KAT7复合物主要由辅助蛋白MEAF6(MYST/esa1 associated factor 6)、ING4(inhibitor of growth family member 4)或ING5(inhibitor of growth family member 5)以及2种与染色质结合的支架蛋白JADE(jade family PHD finger)和BRPF(bromodomain and PHD finger)组成[19]。支架蛋白是KAT7和辅助蛋白的连接点,其相互作用可以调节HAT复合物的底物特异性和活性;KAT7通过与不同的支架蛋白结合形成多种HAT复合物,调控不同组蛋白的乙酰化。例如KAT7-JADE复合物可以使染色质中的组蛋白H3和H4乙酰化,但对H4更有特异性,而KAT7-BRPF复合物则更倾向于乙酰化组蛋白H3[20-21]。
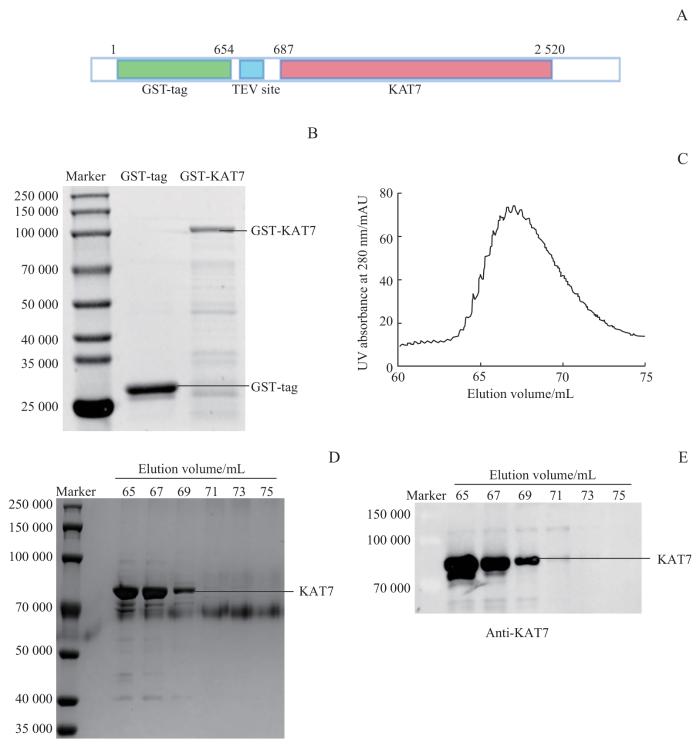
Note: A. KAT7 recombinant protein expression plasmid construction. GST-KAT7 recombinant protein expression plasmid containing GST protein tag with TEV protease restriction site was constructed by using PGEX-4T1 plasmid. B. Expression and purification of GST-KAT7 protein. The recombinant protein GST-KAT7 was expressed by Escherichia coli. BL21 (DE3), and the protein was purified by using GST affinity chromatography, while expressing the GST tag alone was taken as a negative control. C. Purification of KAT7 protein using volume exclusion chromatography. The KAT7 protein after excision of the GST tag was purified by using volume exclusion chromatography HiLoad 16/600 Superdex 75 pg, and peak samples were collected. D. KAT7 samples were separated by SDS-PAGE and determined by Coomassie brilliant blue staining, and the gels were photographed by using visualizer. E. Western blotting assay was performed on KAT7 samples to determine the purity and specificity.
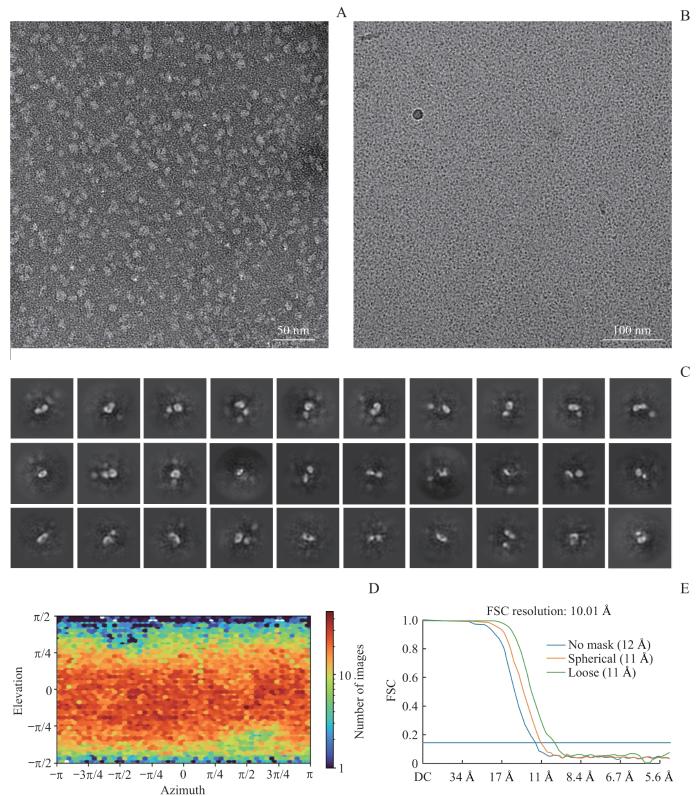
Note: A. The representative negative-staining electron micrograph image of KAT7. B. The representative cryo-electron microscopy micrograph image of KAT7. C. Representative two-dimensional class averages obtained from electron microscopy particles of KAT7. D. Angular distributions of particles for the final reconstruction of KAT7. E. The FSC curves corresponding to the KAT7 three-dimensional structural model. The DC term is 0 Hz and is equivalent to the average of all the samples in the window.
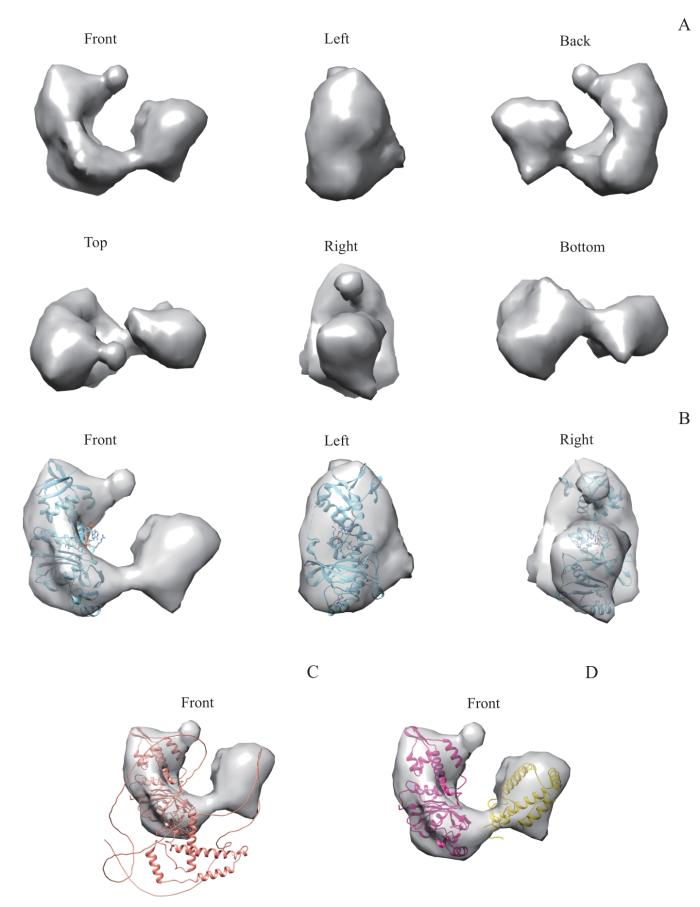
Note: A. Surface view of the electron micrograph density map of KAT7 shown in six orthogonal views. The three-dimensional model size of KAT7 is denoted on the left. B. The crystal structure of MYST motif (5GK9) was docked into corresponding mass of KAT7 shown in three orthogonal views. C. The AlphaFold prediction structure of KAT7 was docked into full-length KAT7 model. D. The N-terminal structural domain (amino acid residues 183‒335, yellow) and MYST domain (amino acid residues 336‒611, red) of the AlphaFold prediction model were docked to the full-length KAT7 model separately.
SHEN Jianfeng designed and planned this study; CAO Qin directed this study; ZHENG Guopei completed all experiments and wrote the paper. All authors have read and agreed to the submission of the manuscript.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
KIM M S, CHO H I, PARK S H, et al. The histone acetyltransferase Myst2 regulates Nanog expression, and is involved in maintaining pluripotency and self-renewal of embryonic stem cells[J]. FEBS Lett, 2015, 589(8): 941-950.
WU Q S, HUANG Q Y, GUAN H L, et al. Comprehensive genome-wide analysis of histone acetylation genes in roses and expression analyses in response to heat stress[J]. Genes, 2022, 13(6): 980.
MARMORSTEIN R, ZHOU M M. Writers and readers of histone acetylation: structure, mechanism, and inhibition[J]. Cold Spring Harb Perspect Biol, 2014, 6(7): a018762.
XING G F, JIN M S, QU R F, et al. Genome-wide investigation of histone acetyltransferase gene family and its responses to biotic and abiotic stress in foxtail millet (Setaria italica[L.]P. Beauv)[J]. BMC Plant Biol, 2022, 22(1): 292.
WANG W, ZHENG Y X, SUN S H, et al. A genome-wide CRISPR-based screen identifies KAT7 as a driver of cellular senescence[J]. Sci Transl Med, 2021, 13(575): eabd2655.
HEINLEIN M, GANDOLFO L C, ZHAO K L, et al. The acetyltransferase KAT7 is required for thymic epithelial cell expansion, expression of AIRE target genes, and thymic tolerance[J]. Sci Immunol, 2022, 7(67): eabb6032.
HAN Z, WU H, KIM S, et al. Revealing the protein propionylation activity of the histone acetyltransferase MOF (males absent on the first)[J]. J Biol Chem, 2018, 293(9): 3410-3420.
YAN M S, TURGEON P J, MAN H S J, et al. Histone acetyltransferase 7 (KAT7)-dependent intragenic histone acetylation regulates endothelial cell gene regulation[J]. J Biol Chem, 2018, 293(12): 4381-4402.
NEWMAN D M, VOSS A K, THOMAS T, et al. Essential role for the histone acetyltransferase KAT7 in T cell development, fitness, and survival[J]. J Leukoc Biol, 2017, 101(4): 887-892.
SAKSOUK N, AVVAKUMOV N, CHAMPAGNE K S, et al. HBO1 HAT complexes target chromatin throughout gene coding regions via multiple PHD finger interactions with histone H3 tail[J]. Mol Cell, 2009, 33(2): 257-265.
MIOTTO B, STRUHL K. JNK1 phosphorylation of Cdt1 inhibits recruitment of HBO1 histone acetylase and blocks replication licensing in response to stress[J]. Mol Cell, 2011, 44(1): 62-71.
TAO Y, ZHONG C, ZHU J J, et al. Structural and mechanistic insights into regulation of HBO1 histone acetyltransferase activity by BRPF2[J]. Nucleic Acids Res, 2017, 45(10): 5707-5719.
AU Y Z, GU M X, DE BRAEKELEER E, et al. KAT7 is a genetic vulnerability of acute myeloid leukemias driven by MLL rearrangements[J]. Leukemia, 2021, 35(4): 1012-1022.
MCCULLOUGH C E, MARMORSTEIN R. Molecular basis for histone acetyltransferase regulation by binding partners, associated domains, and autoacetylation[J]. ACS Chem Biol, 2016, 11(3): 632-642.
YAN K Z, YOU L Y, DEGERNY C, et al. The chromatin regulator BRPF3 preferentially activates the HBO1 acetyltransferase but is dispensable for mouse development and survival[J]. J Biol Chem, 2016, 291(6): 2647-2663.
KUEH A J, BERGAMASCO M I, QUAGLIERI A, et al. Stem cell plasticity, acetylation of H3K14, and de novo gene activation rely on KAT7[J]. Cell Rep, 2023, 42(1): 111980.
XIAO Y H, LI W J, YANG H, et al. HBO1 is a versatile histone acyltransferase critical for promoter histone acylations[J]. Nucleic Acids Res, 2021, 49(14): 8037-8059.
IIZUKA M, SARMENTO O F, SEKIYA T, et al. Hbo1 links p53-dependent stress signaling to DNA replication licensing[J]. Mol Cell Biol, 2008, 28(1): 140-153.
LAN R F, WANG Q Q. Deciphering structure, function and mechanism of lysine acetyltransferase HBO1 in protein acetylation, transcription regulation, DNA replication and its oncogenic properties in cancer[J]. Cell Mol Life Sci, 2020, 77(4): 637-649.
... 人源性KAT7具有611个氨基酸残基,主要由2个结构域组成:N端结构域包含1个短的锌指DNA结合域,它主要与MCM2(minichromosome maintenance complex component 2)和ORC1(origin recognition complex subunit 1)相互作用;C端为MYST乙酰基转移酶结构域,主要负责结合乙酰辅酶A并催化对应的乙酰化反应[16].MYST结构域是MYST家族共有的乙酰转移酶结构域,其在多种HAT蛋白如MYST1(MOF/KAT8)、MYST2(HBO1/KAT7)和MYST3(MoZ/KAT6A)中高度保守[17].HAT通常与支架蛋白和其他辅助蛋白相互作用,形成HAT复合物并发挥功能[18].已经鉴定的KAT7复合物主要由辅助蛋白MEAF6(MYST/esa1 associated factor 6)、ING4(inhibitor of growth family member 4)或ING5(inhibitor of growth family member 5)以及2种与染色质结合的支架蛋白JADE(jade family PHD finger)和BRPF(bromodomain and PHD finger)组成[19].支架蛋白是KAT7和辅助蛋白的连接点,其相互作用可以调节HAT复合物的底物特异性和活性;KAT7通过与不同的支架蛋白结合形成多种HAT复合物,调控不同组蛋白的乙酰化.例如KAT7-JADE复合物可以使染色质中的组蛋白H3和H4乙酰化,但对H4更有特异性,而KAT7-BRPF复合物则更倾向于乙酰化组蛋白H3[20-21]. ...
... 人源性KAT7具有611个氨基酸残基,主要由2个结构域组成:N端结构域包含1个短的锌指DNA结合域,它主要与MCM2(minichromosome maintenance complex component 2)和ORC1(origin recognition complex subunit 1)相互作用;C端为MYST乙酰基转移酶结构域,主要负责结合乙酰辅酶A并催化对应的乙酰化反应[16].MYST结构域是MYST家族共有的乙酰转移酶结构域,其在多种HAT蛋白如MYST1(MOF/KAT8)、MYST2(HBO1/KAT7)和MYST3(MoZ/KAT6A)中高度保守[17].HAT通常与支架蛋白和其他辅助蛋白相互作用,形成HAT复合物并发挥功能[18].已经鉴定的KAT7复合物主要由辅助蛋白MEAF6(MYST/esa1 associated factor 6)、ING4(inhibitor of growth family member 4)或ING5(inhibitor of growth family member 5)以及2种与染色质结合的支架蛋白JADE(jade family PHD finger)和BRPF(bromodomain and PHD finger)组成[19].支架蛋白是KAT7和辅助蛋白的连接点,其相互作用可以调节HAT复合物的底物特异性和活性;KAT7通过与不同的支架蛋白结合形成多种HAT复合物,调控不同组蛋白的乙酰化.例如KAT7-JADE复合物可以使染色质中的组蛋白H3和H4乙酰化,但对H4更有特异性,而KAT7-BRPF复合物则更倾向于乙酰化组蛋白H3[20-21]. ...
1
... 人源性KAT7具有611个氨基酸残基,主要由2个结构域组成:N端结构域包含1个短的锌指DNA结合域,它主要与MCM2(minichromosome maintenance complex component 2)和ORC1(origin recognition complex subunit 1)相互作用;C端为MYST乙酰基转移酶结构域,主要负责结合乙酰辅酶A并催化对应的乙酰化反应[16].MYST结构域是MYST家族共有的乙酰转移酶结构域,其在多种HAT蛋白如MYST1(MOF/KAT8)、MYST2(HBO1/KAT7)和MYST3(MoZ/KAT6A)中高度保守[17].HAT通常与支架蛋白和其他辅助蛋白相互作用,形成HAT复合物并发挥功能[18].已经鉴定的KAT7复合物主要由辅助蛋白MEAF6(MYST/esa1 associated factor 6)、ING4(inhibitor of growth family member 4)或ING5(inhibitor of growth family member 5)以及2种与染色质结合的支架蛋白JADE(jade family PHD finger)和BRPF(bromodomain and PHD finger)组成[19].支架蛋白是KAT7和辅助蛋白的连接点,其相互作用可以调节HAT复合物的底物特异性和活性;KAT7通过与不同的支架蛋白结合形成多种HAT复合物,调控不同组蛋白的乙酰化.例如KAT7-JADE复合物可以使染色质中的组蛋白H3和H4乙酰化,但对H4更有特异性,而KAT7-BRPF复合物则更倾向于乙酰化组蛋白H3[20-21]. ...
1
... 人源性KAT7具有611个氨基酸残基,主要由2个结构域组成:N端结构域包含1个短的锌指DNA结合域,它主要与MCM2(minichromosome maintenance complex component 2)和ORC1(origin recognition complex subunit 1)相互作用;C端为MYST乙酰基转移酶结构域,主要负责结合乙酰辅酶A并催化对应的乙酰化反应[16].MYST结构域是MYST家族共有的乙酰转移酶结构域,其在多种HAT蛋白如MYST1(MOF/KAT8)、MYST2(HBO1/KAT7)和MYST3(MoZ/KAT6A)中高度保守[17].HAT通常与支架蛋白和其他辅助蛋白相互作用,形成HAT复合物并发挥功能[18].已经鉴定的KAT7复合物主要由辅助蛋白MEAF6(MYST/esa1 associated factor 6)、ING4(inhibitor of growth family member 4)或ING5(inhibitor of growth family member 5)以及2种与染色质结合的支架蛋白JADE(jade family PHD finger)和BRPF(bromodomain and PHD finger)组成[19].支架蛋白是KAT7和辅助蛋白的连接点,其相互作用可以调节HAT复合物的底物特异性和活性;KAT7通过与不同的支架蛋白结合形成多种HAT复合物,调控不同组蛋白的乙酰化.例如KAT7-JADE复合物可以使染色质中的组蛋白H3和H4乙酰化,但对H4更有特异性,而KAT7-BRPF复合物则更倾向于乙酰化组蛋白H3[20-21]. ...
1
... 人源性KAT7具有611个氨基酸残基,主要由2个结构域组成:N端结构域包含1个短的锌指DNA结合域,它主要与MCM2(minichromosome maintenance complex component 2)和ORC1(origin recognition complex subunit 1)相互作用;C端为MYST乙酰基转移酶结构域,主要负责结合乙酰辅酶A并催化对应的乙酰化反应[16].MYST结构域是MYST家族共有的乙酰转移酶结构域,其在多种HAT蛋白如MYST1(MOF/KAT8)、MYST2(HBO1/KAT7)和MYST3(MoZ/KAT6A)中高度保守[17].HAT通常与支架蛋白和其他辅助蛋白相互作用,形成HAT复合物并发挥功能[18].已经鉴定的KAT7复合物主要由辅助蛋白MEAF6(MYST/esa1 associated factor 6)、ING4(inhibitor of growth family member 4)或ING5(inhibitor of growth family member 5)以及2种与染色质结合的支架蛋白JADE(jade family PHD finger)和BRPF(bromodomain and PHD finger)组成[19].支架蛋白是KAT7和辅助蛋白的连接点,其相互作用可以调节HAT复合物的底物特异性和活性;KAT7通过与不同的支架蛋白结合形成多种HAT复合物,调控不同组蛋白的乙酰化.例如KAT7-JADE复合物可以使染色质中的组蛋白H3和H4乙酰化,但对H4更有特异性,而KAT7-BRPF复合物则更倾向于乙酰化组蛋白H3[20-21]. ...
1
... 人源性KAT7具有611个氨基酸残基,主要由2个结构域组成:N端结构域包含1个短的锌指DNA结合域,它主要与MCM2(minichromosome maintenance complex component 2)和ORC1(origin recognition complex subunit 1)相互作用;C端为MYST乙酰基转移酶结构域,主要负责结合乙酰辅酶A并催化对应的乙酰化反应[16].MYST结构域是MYST家族共有的乙酰转移酶结构域,其在多种HAT蛋白如MYST1(MOF/KAT8)、MYST2(HBO1/KAT7)和MYST3(MoZ/KAT6A)中高度保守[17].HAT通常与支架蛋白和其他辅助蛋白相互作用,形成HAT复合物并发挥功能[18].已经鉴定的KAT7复合物主要由辅助蛋白MEAF6(MYST/esa1 associated factor 6)、ING4(inhibitor of growth family member 4)或ING5(inhibitor of growth family member 5)以及2种与染色质结合的支架蛋白JADE(jade family PHD finger)和BRPF(bromodomain and PHD finger)组成[19].支架蛋白是KAT7和辅助蛋白的连接点,其相互作用可以调节HAT复合物的底物特异性和活性;KAT7通过与不同的支架蛋白结合形成多种HAT复合物,调控不同组蛋白的乙酰化.例如KAT7-JADE复合物可以使染色质中的组蛋白H3和H4乙酰化,但对H4更有特异性,而KAT7-BRPF复合物则更倾向于乙酰化组蛋白H3[20-21]. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}