

铁死亡(ferroptosis)是一种铁依赖和磷脂过氧化驱动的程序性细胞死亡[1],在缺血再灌注损伤、肿瘤耐药和退行性疾病中均发挥重要作用[2-4]。铁死亡的主要特征是铁稳态的失调以及脂质过氧化物的累积[5]。研究表明,铁死亡诱导剂可以引起关键基因表达的改变,从而诱导细胞脂质过氧化物的累积及铁死亡的发生。其中,酰基辅酶A合成酶长链家族成员4(acyl-CoA synthetase long-chain family member 4,ACSL4)能够将多不饱和脂肪酸(polyunsaturated fatty acid,PUFA)添加到磷脂上,这对膜磷脂过氧化物的累积和铁死亡的发生至关重要[6]。谷胱甘肽过氧化物酶4(glutathione peroxidase 4,GPX4)则通过利用谷胱甘肽(glutathione,GSH)提供的电子将磷脂和胆固醇氢过氧化物还原成相应的醇,从而避免细胞内脂质过氧化物的过度累积,抑制铁死亡的发生[7]。然而,这些关键分子的翻译后修饰是否在铁死亡中发挥作用还有待研究。
蛋白质小分子泛素相关修饰物蛋白(small ubiquitin-related modifier protein,SUMO)修饰是一种可逆的翻译后修饰过程,在各种修饰酶的作用下保持着动态平衡,能改变蛋白质的稳定性、功能或定位,影响细胞内诸多生命活动,参与细胞生理与病理过程的调控。SUMO化的重要性已经在多种细胞死亡途径中得到证明。有证据[8]表明,SUMO化可能与铁死亡有关,但其具体机制尚未报道。kelch样ECH关联蛋白1(kelch-like ECH-associated protein 1,KEAP1)/核因子E2相关因子2(nuclear factor erythroid 2-related factor 2,NRF2)[9-10]、P53[11]、苄氯素1(beclin 1,BECN1)[12]和神经纤维蛋白2(neurofibromin 2,NF2)/YY1相关蛋白1(YY1-associated protein 1,YY1AP1)[13]等一系列SUMO靶蛋白会调控细胞铁死亡。例如,SUMO化修饰能够提高NRF2清除活性氧(reactive oxygen species,ROS)的活性,进而影响肝癌细胞脂质过氧化水平[9-10]。这些研究提示蛋白质SUMO化与铁死亡之间的关系对相关疾病的发生发展具有一定的意义,可能可以作为治疗的靶点。
SUMO特异性蛋白酶(SUMO specific peptidase,SENP)是一类半胱氨酸蛋白酶,可促进SUMO蛋白的去偶联,参与维持SUMO化和非SUMO化蛋白之间的平衡。以SENP为靶点是治疗某些疾病的新策略。作为其中的一员,SENP1在肿瘤中主要发挥促癌作用[14]。研究[15]报道,SENP1可以通过使低氧诱导因子-1α(hypoxia inducible factor-1α,HIF-1α)去SUMO化增加其蛋白稳定性,从而增强肝癌细胞的细胞干性,促进肝癌的发生。此外,SENP1可以调节β-联蛋白(β-catenin)的去SUMO化进而促进乳腺癌的进展[16]。近期有研究[17]发现,SENP1对受体相互作用蛋白激酶1(receptor-interacting protein kinase 1,RIPK1)的去SUMO化修饰能够抑制RIPK1驱动的细胞凋亡和炎症。然而,SENP1在铁死亡中是否发挥作用以及作用的机制尚未见报道。本研究旨在探讨SENP1在铁死亡中的潜在作用及其可能的机制。
1 材料与方法
1.1 细胞和质粒
人纤维肉瘤细胞系HT1080由上海交通大学医学院杨文教授惠赠,鼠纤维肉瘤细胞系MCA-205购自上海恒雅生物科技有限公司,人胚胎肾细胞株293T购自美国模式菌种保藏中心(American Type Culture Collection,ATCC)。所有细胞均用DMEM高糖完全培养基[含10%胎牛血清(fetal bovine serum,FBS)和1%双抗],于37 ℃、含5% CO2培养箱中培养。
pcDNA4-Flag-Vector质粒为本实验室库存,Flag-SENP1质粒由上海交通大学医学院程金科教授惠赠。
1.2 主要试剂和仪器
DMEM高糖液体培养基、0.25%胰蛋白酶、细胞冻存液(上海源培生物科技股份有限公司),FBS、opti-MEM培养基(Gibco,美国),青/链霉素双抗(Cytiva,美国),PBS、β-肌动蛋白(β-actin)抗体(武汉赛维尔生物科技有限公司),ACSL4抗体(Santa Cruz Biotechnology,美国),GPX4抗体、2×SYBR Green Fast qPCR Mix荧光定量试剂盒、SENP1抗体(武汉爱博泰克生物科技有限公司),BCA蛋白定量试剂(Thermo,美国),铁死亡诱导剂RSL3(RAS-selective lethal 3)和erastin(MedChemExpress,美国),二甲基亚砜(DMSO)、Flag抗体、Anti-Flag M2 Beads(Sigma-Aldrich,德国),质粒中提试剂盒(Qiagen,德国),RNA抽提试剂盒、反转录试剂盒、抗体稀释液、胶回收试剂盒、LB培养基(南京诺唯赞生物科技有限公司),辣根过氧化酶(HRP)-羊抗兔/鼠二抗(武汉三鹰生物科技有限公司),线性化聚乙烯亚胺(PEI)[翌圣生物科技(上海)股份有限公司]。
Nanodrop 2000微量紫外分光光度计(Thermo Fisher,美国),细胞计数仪(Counterstar,中国)、蛋白免疫印迹电泳仪(Bio-Rad,美国),正置光学显微镜(Nikon,日本),Amersham ImageQuant 800蛋白印迹成像系统(Cytiva,美国),qTOWER 3G荧光定量PCR仪(耶拿,德国)。
1.3 生物信息学分析
使用肿瘤基因组图谱(The Cancer Genome Atlas,TCGA;
1.4 实时荧光定量PCR
使用Buffer RL进行消化、裂解,涡旋振荡直至无明显细胞团。使用RNA抽提试剂盒提取RNA,定量后使用反转录试剂盒将mRNA反转录为cDNA,稀释20倍后作为模板。采用2×SYBR Green Fast qPCR Mix荧光定量试剂盒。引物由苏州金唯智生物科技有限公司合成,序列见表1。实时荧光定量PCR(RT-qPCR)体系配置,每孔10 µL(含2×SYBR Green Fast qPCR Mix 5 µL、正向引物0.5 µL、反向引物0.5 µL、模板1 µL和ddH2O 3 µL),每个样品设置3个复孔。点样于384孔板中,贴膜,4 ℃下1 500×g平转离心5 min。荧光定量PCR仪扩增。PCR扩增程序为:95 ℃ 10 min;95 ℃ 15 s,60 ℃ 1 min,共40个循环。
表1 RT-qPCR引物序列
Tab 1
| Gene | Forward primer (5′→3′) | Reverse primer (5′→3′) |
|---|---|---|
| SENP1 | CCAGATTGAAGAACAGAA | GACAACAGTAACAGGAAT |
| GAPDH | ACATCGCTCAGACACCATG | TGTAGTTGAGGTCAATGAAGGG |
| ACSL4 | CATCCCTGGAGCAGATACTCT | TCACTTAGGATTTCCCTGGTCC |
| GPX4 | GAGGCAAGACCGAAGTAAACTAC | CCGAACTGGTTACACGGGAA |
1.5 Western blotting
收集细胞样品,裂解变性,用10%十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE),先80 V,20 min,随后150 V,60 min。转膜设置80 V,2 h,将蛋白转移至PVDF膜上。用5%的脱脂牛奶室温摇床封闭30 min,4 ℃摇床上一抗孵育过夜[SENP1抗体(1∶1 000)、β-actin抗体(1∶1 000)、Flag抗体(1∶5 000)、ACSL4抗体(1∶500)、GPX4抗体(1∶1 000)]。收集一抗,用PBST缓冲液洗膜,每次10 min,重复3次。采用化学发光法,用HRP标记的二抗(1∶5 000)室温摇床孵育1 h,将显影液按1∶1配置,均匀滴加在膜上。采用自动曝光,根据结果选择合适的曝光时间和次数。结果用Image Lab Touch软件处理分析。
1.6 免疫沉淀-质谱联用技术
1.6.1 样品准备
准备好生长状态良好的293T细胞,转染前2 h细胞换液。预热opti-MEM和PEI。转染体系:混合物1(500 µL opti-MEM中加入10 µg pcDNA4-Flag-Vector或Flag-SENP1)、混合物2(500 µL opti-MEM中加入40 µL PEI)。两者静置5 min后混合,继续静置15 min,将配置的转染体系加入293T细胞培养液中,6~8 h后换液。样品分为3组,分别为Flag-Vector组、Flag-SENP1组及Flag-SENP1+erastin组。
1.6.2 免疫沉淀实验
转染24 h后收集细胞样品。收集样品前3 h,Flag-SENP1+erastin组加入铁死亡诱导剂erastin(终浓度2.5 μmol/L)。用细胞刮收集细胞样品,4 ℃下700×g离心5 min,预冷PBS洗一遍;加1 mL TAP裂解缓冲液冰上裂解20 min,4 ℃下12 000×g离心10 min,取45 μL上清液作为input。用TAP裂解缓冲液清洗Anti-Flag M2 Beads 3次;700×g离心3 min,用90 μL TAP裂解缓冲液重悬Anti-Flag M2 Beads,每1 mL细胞裂解液样品加入20 μL Anti-Flag M2 Beads重悬液,旋转混合仪上4 ℃孵育过夜;4 ℃下900×g离心3 min,弃上清液;用TAP裂解缓冲液清洗3次,弃上清液;用Flag洗脱缓冲液进行洗脱,4 ℃下900×g离心3 min,重复3次;加入含0.2 μg/mL Flag peptide Flag洗脱缓冲液,在旋转混合仪上4 ℃洗脱2 h;4 ℃下900×g离心3 min,取上清液。
1.6.3 质谱分析
收集3组样品,并进行10% SDS-PAGE检测,切胶收集样品送上海交通大学分析测试中心进行质谱分析。Flag-SENP1+erastin组的质谱检测结果中,去除角蛋白,以及与Flag-SENP1组重复的蛋白,对剩余的结合蛋白根据分值进行排序;分值越高,说明SENP1对该蛋白的富集越多。
1.7 统计学分析
使用GraphPad Prism 8.0软件进行统计学分析。定量资料用x±s表示;2组间比较采用t检验,多组间比较使用方差分析,有差异的再进行两两比较(t检验)。P<0.05表示差异具有统计学意义。所有实验均重复3次。
2 结果
2.1 数据库中 SENP1 与 ACSL4 和 GPX4 表达的相关性
图1
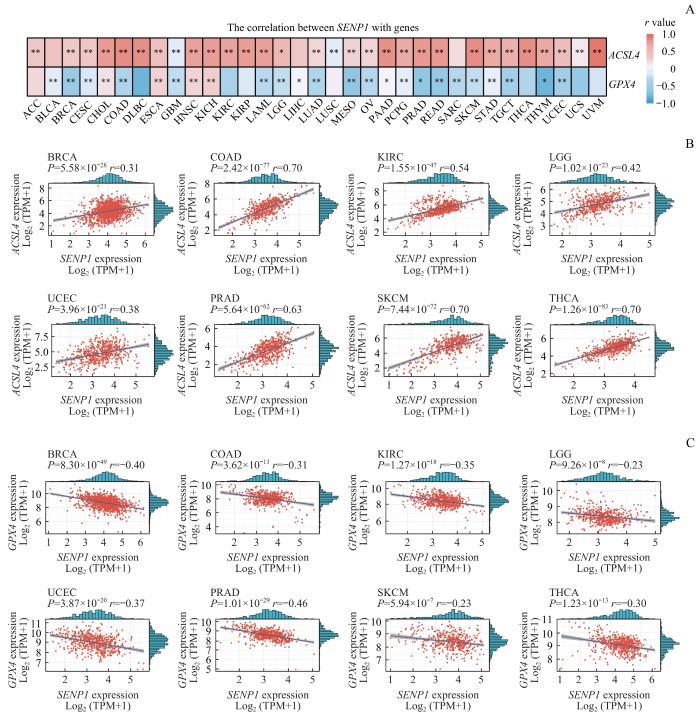
图1
TCGA中 SENP1 和 ACSL4/GPX4 的相关性分析
Note: A. The correlation between SENP1 and ACSL4, as well as GPX4, in the TCGA database. B. Scatter plots of the correlation of SENP1 with ACSL4 in the representative tumour tissues. C. Scatter plots of the correlation of SENP1 with GPX4 in the representative tumour tissues. ACC—adenoid cystic carcinoma; BLCA—bladder urothelial carcinoma; BRCA—breast invasive carcinoma; CESC—cervical squamous cell carcinoma and endocervical adenocarcinoma; CHOL—cholangiocarcinoma; COAD—colon adenocarcinoma; DLBC—lymphoid neoplasm diffuse large B-cell lymphoma; ESCA—esophageal carcinoma; GBM—glioblastoma multiforme; HNSC—head and neck squamous cell carcinoma; KICH—kidney chromophobe; KIRC—kidney renal clear cell carcinoma; KIRP—kidney renal papillary cell carcinoma; LAML—acute myeloid leukemia; LGG—low-grade glioma; LIHC—liver hepatocellular carcinoma; LUAD—lung adenocarcinoma; LUSC—lung squamous cell carcinoma; MESO—mesothelioma; OV—ovarian serous cystadenocarcinoma; PAAD—pancreatic adenocarcinoma; PCPG—pheochromocytoma and Paraganglioma; PRAD—prostate adenocarcinoma; READ—rectum adenocarcinoma; SARC—sarcoma; SKCM—skin cutaneous melanoma; STAD—stomach adenocarcinoma; TGCT—testicular germ cell tumors; THCA—thyroid carcinoma; THYM—thymoma; UCEC—uterine corpus endometrial carcinoma; UCS—uterine carcinosarcom; UVM—uveal melanoma; TPM—transcripts per million.
Fig 1
Correlation analysis of SENP1 with ACSL4/GPX4 in the TCGA database
2.2 铁死亡过程中SENP1表达水平下调
为了验证SENP1在铁死亡中的作用,使用GPX4抑制剂RSL3分别诱导人纤维肉瘤细胞系HT1080、鼠纤维肉瘤细胞系MCA-205、人胚胎肾细胞株293T铁死亡,检测SENP1 mRNA和蛋白的表达水平(图2)。RT-qPCR结果显示,HT1080细胞中SENP1 mRNA水平在处理2 h左右开始降低,4~10 h下降显著。Western blotting结果显示,HT1080细胞中SENP1蛋白的表达水平在6 h之后明显下调。用5 μmol/L和10 μmol/L RSL3处理MCA-205细胞5~10 h,SENP1蛋白水平明显下调。我们在293T细胞中发现了类似的结果,使用0.25 μmol/L RSL3诱导293T细胞铁死亡。RT-qPCR结果显示,293T细胞中SENP1的mRNA水平在2 h左右开始降低,4~8 h下降显著;Western blotting结果显示,293T细胞中SENP1蛋白水平在4 h之后显著下调。
图2
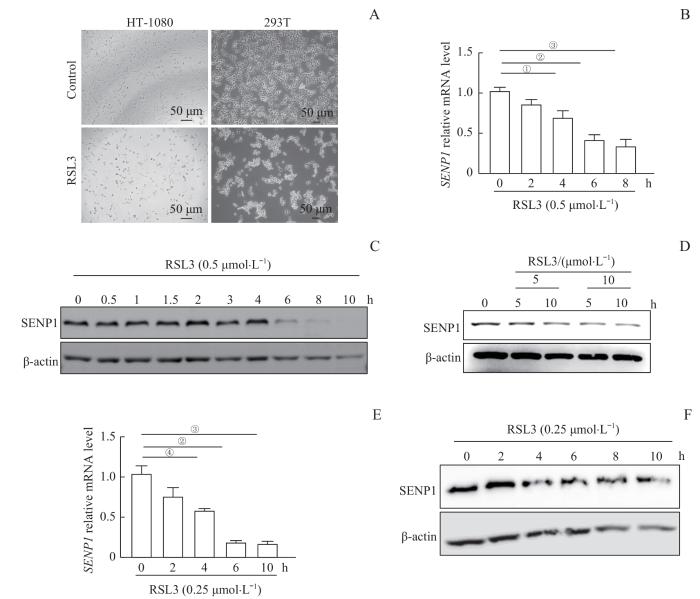
图2
铁死亡诱导过程中 SENP1 的mRNA和蛋白表达水平
Note: A. Bright-field images of HT1080 and 293T cells treated with RSL3 (×20, scale bar=50 μm). B. mRNA levels of SENP1 in HT1080 cells treated with 0.5 μmol/L RSL3 for the indicated time, as detected by RT-qPCR. C. Protein levels of SENP1 in HT1080 cells treated with 0.5 μmol/L RSL3 for the indicated time, as detected by Western blotting. D. Protein levels of SENP1 in MCA-205 cells treated with 5 or 10 μmol/L RSL3 for 5 or 10 h, as detected by Western blotting. E. mRNA levels of SENP1 in 293T cells treated with 0.25 μmol/L RSL3 for the indicated time, as detected by RT-qPCR. F. Protein levels of SENP1 in 293T cells treated with 0.25 μmol/L RSL3 for the indicated time, as detected by Western blotting. ①P=0.025, ②P=0.004, ③P=0.001, ④P=0.034.
Fig 2
mRNA and protein expression levels of SENP1 during ferroptosis induction
2.3 免疫沉淀-质谱联用技术鉴定SENP1相互作用蛋白
接下来我们通过免疫沉淀-质谱联用(immunoprecipitation-mass spectrometry,IP-MS)技术初步探讨SENP1在铁死亡发生过程中的相互作用蛋白(图3)。首先,在293T细胞中转染pcDNA4-Flag-Vector或Flag-SENP1过表达质粒,收蛋白前3 h使用2.5 μmol/L erastin处理293T细胞。Western blotting结果显示,Flag-SENP1在293T细胞中成功表达。SDS-PAGE后将免疫沉淀样品送质谱分析,鉴定结果显示,在erastin诱导情况下,铁死亡发生过程中SENP1可富集SUMO分子,提示在铁死亡发生过程中可能有蛋白发生了去SUMO化。
图3
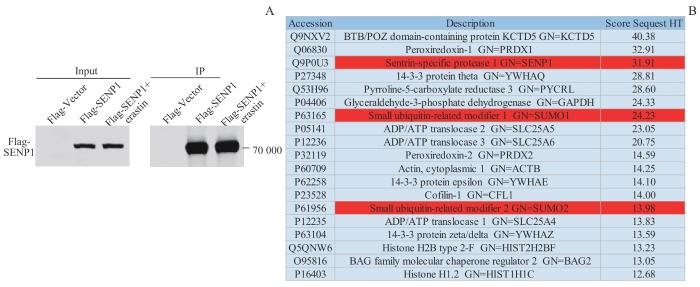
图3
IP-MS技术分析SENP1在铁死亡发生过程中相互作用蛋白
Note: A. Expression levels of Flag-SENP1 in 293T cells, detected by Western blotting. B. Identification results of SENP1 by IP-MS, with analysis and screening the top candidates that may interact with SENP1.
Fig 3
Interacting proteins of SENP1 detected by IP-MS during ferroptosis
2.4 过表达SENP1后ACSL4蛋白水平上调
随后,我们检测转染Flag-Vector或Flag-SENP1的293T细胞中铁死亡相关基因的表达变化(图4)。RT-qPCR检测结果显示,Flag-SENP1组SENP1的mRNA水平相较于Flag-Vector组显著升高。Western blotting结果显示,Flag-SENP1蛋白在Flag-SENP1组中成功表达。同时,与Flag-Vector组相比,Flag-SENP1组293T细胞中ACSL4蛋白表达水平上调,而GPX4蛋白水平未见明显变化。RT-qPCR结果显示,过表达SENP1后,ACSL4和GPX4在mRNA水平表达无显著改变。以上结果提示SENP1可能通过调控ACSL4的蛋白稳定性来影响铁死亡的发生。
图4
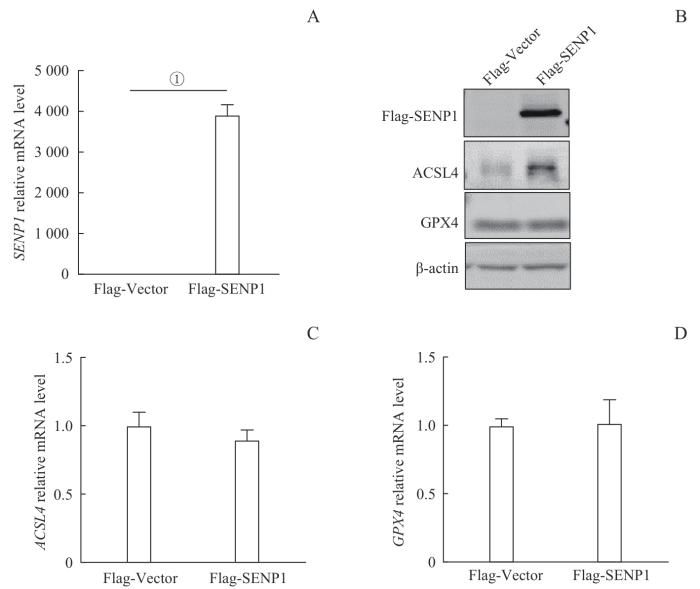
图4
检测过表达SENP1后铁死亡相关基因的表达情况
Note: A. SENP1 mRNA expression in the transfected 293T cells, detected by RT-qPCR. B. Expression of Flag-SENP1/ACSL4/GPX4 in the transfected 293T cells, detected by Western blotting. C. ACSL4 mRNA expression in the transfected 293T cells, detected by RT-qPCR. D. GPX4 mRNA expression in the transfected 293T cells, detected by RT-qPCR. ①P<0.001.
Fig 4
Expression of ferroptosis-related genes after overexpression of SENP1
3 讨论
真核生物中蛋白质的SUMO化修饰是一个可逆的过程,在各种蛋白酶的作用下保持着动态平衡。SUMO主要包括4个亚型,即SUMO1~4,它们在从酵母到人的真核生物中高度保守。蛋白质的SUMO化修饰通常由E1激活酶、E2结合酶和E3连接酶相继完成,去SUMO化过程则由具有木瓜蛋白酶折叠结构的半胱氨酸水解酶SENP所介导。迄今为止,在酵母中发现了2种SENP——ULP1(SUMO protease ULP1,ULP1)和ULP2;人类基因组编码6种SENP,分别为SENP1~3、SENP5~7[18]。SUMO化修饰几乎存在于所有的真核生物中,它可以改变蛋白的许多特征,例如转录活性、稳定性、结构、功能、细胞内亚定位以及与其他蛋白的相互作用等[19],对于维持多种细胞生物学过程、基因表达及信号转导的调控非常重要。
于是,我们在HT1080、MCA-205和293T细胞中用RSL3诱导铁死亡,结果发现SENP1的表达水平有明显的降低趋势。为了探究SENP1在铁死亡发生的过程中发挥着何种作用,我们通过IP-MS试图寻找SENP1相互作用蛋白,鉴定结果显示,诱导铁死亡发生时,SENP1可富集SUMO分子,提示在铁死亡发生的过程中可能有蛋白发生了去SUMO化。那么SENP1可能影响铁死亡的哪条通路呢?已有研究表明,铁死亡的发生可以被脂质、氨基酸、铁等几种代谢途径调控。铁死亡最经典的防御途径为溶质载体家族7成员11(solute carrier family 7 member 11,SLC7A11)/GSH/GPX4通路,其他还包括定位在细胞膜上的铁死亡抑制蛋白1[20](ferroptosis suppressor protein 1,FSP1)和线粒体中的二氢乳清酸脱氢酶(dihydroorotate dehydrogenase,DHODH)[21]等。细胞内铁代谢的调控主要是通过转铁蛋白和转铁蛋白受体从细胞外摄入三价铁,三价铁在还原酶的作用下转变为二价铁,而细胞内过量的亚铁离子会促进芬顿反应和脂质过氧化,导致铁死亡的发生,因此转铁蛋白及其受体对铁死亡也至关重要[22]。另一个铁死亡关键蛋白是ACSL4。ACSL4作为长链脂酰辅酶A合成酶家族成员,能将长链多不饱和脂肪酸活化,参与膜磷脂的合成,而这些长链多不饱和脂肪酸常被氧化,尤其是在RSL3等药物的诱导下。已有研究[23]表明ACSL4可作为胶质瘤中重要的铁死亡标志物。当然,铁死亡调控通路还远不止这些。
因此,接下来我们在293T细胞中转染Flag-SENP1过表达质粒,发现过表达SENP1后铁死亡标志蛋白ACSL4蛋白水平表达上调。结果提示,SENP1可能通过调控ACSL4的蛋白稳定性影响铁死亡的发生。但是,SENP1是否调控ACSL4的去SUMO化修饰,以及去SUMO化修饰的位点还有待在日后的工作中进一步探索。以上结果提示,SENP1可促进铁死亡发生。值得注意的是,RSL3诱导的铁死亡过程中SENP1表达水平下调,我们推测铁死亡过程触发了细胞内的保护机制,通过下调SENP1基因的表达以抵抗铁死亡的发生,但是其具体机制有待明确。在之后的研究中,我们将会构建SENP1基因条件性敲除小鼠,从而通过体内实验明确SENP1基因对铁死亡的影响,同时完善相关体外实验,并明确铁死亡过程中SENP1基因表达下调及其调控铁死亡的机制。
总之,本研究从蛋白质SUMO化的角度出发,发现SENP1基因在铁死亡过程中表达下调,并可能通过调控铁死亡相关蛋白ACSL4的稳定性对铁死亡发挥潜在的作用。这为阐明铁死亡的分子机制提供了新的线索和思路。
作者贡献声明
谢滨参与实验设计、实验实施、数据分析、论文写作与修改;白蒙、吴妍和沃璐璐参与前期实验设计与实施;黄莺参与指导论文写作与修改;张晶全程指导课题开展、论文写作与修改。所有作者均阅读并同意最终稿件的提交。
AUTHOR's CONTRIBUTIONS
XIE Bin participated in experiment design, experiment implementation, data analysis, paper writing and revision. BAI Meng, WU Yan and WO Lulu participated in the preliminary experimental design and implementation. HUANG Ying participated in guiding the writing and revision of the paper. ZHANG Jing supervised the whole project development, paper writing and revision. All the authors have read the final version of paper and consented to its submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclare no relevant conflict of interests.
参考文献

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}