目的·通过单细胞核转录组分析,探讨多发性硬化症(multiple sclerosis,MS)中小胶质细胞在灰质与白质的基因差异性表达及其在疾病进展中的作用,鉴定疾病相关的关键转录调控网络。方法·从基因表达数据库(Gene Expression Omnibus,GEO)中获取MS和对照冷冻人脑组织样本单细胞核转录组测序(single nucleus RNA sequencing,snRNA-seq)数据。使用R软件和Seurat软件等,利用特定的细胞标志物对数据进行细胞类型的鉴定。从鉴定的细胞群中提取小胶质细胞,根据其解剖来源将其分为灰质和白质小胶质细胞;利用降维聚类技术,获得具有差异性的小胶质细胞亚群。使用Seurat分析得到MS组与对照组在亚群层面上的差异表达基因(differentially expressed genes,DEGs)。对DEGs进行基因本体论(Gene Ontology,GO)分析与京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)分析,进一步探究这些差异的生物学意义。使用Monocle3进行拟时序分析,研究疾病进展中的细胞亚群动态变化。使用单细胞调控网络推理和聚类(single cell regulatory network inference and clustering,SCENIC)方法分析转录因子(transcription factor,TF)调控网络,寻找可能参与MS调控的关键转录调控网络。结果·对数据进行质量控制后共保留了149 062个细胞核。对snRNA-seq数据进行降维聚类分析后,以DOCK8、CSF1R、P2RY12、CD74作为小胶质细胞的关键标志物鉴定得到了12 238个小胶质细胞。GO和KEGG分析结果表明,灰质小胶质细胞在疾病过程中内吞作用、离子稳态、脂质定位等功能下调,白质小胶质细胞在疾病过程中蛋白质折叠、细胞质翻译、温度刺激响应等功能上调。SCENIC分析显示MS疾病中FLI1、MITF、FOXP1等TF的表达上调。结论·小胶质细胞在MS的发展中具有重要作用,白质小胶质细胞受到MS的影响比灰质小胶质细胞更为明显。FLI1、MITF、FOXP1等是参与MS调控的关键TF,这些转录调控网络在疾病调控中发挥核心作用。
关键词:多发性硬化症
;
小胶质细胞
;
单细胞核转录组
;
富集分析
;
转录调控网络
Abstract
Objective ·To investigate the differential gene expression of microglia in the gray and white matter of multiple sclerosis (MS) using single-nucleus transcriptomic analysis, aiming to explore their roles in disease progression, and identify key transcriptional regulatory networks associated with the disease. Methods ·snRNA-seq data of frozen human brain tissue samples from MS patients and control individuals were obtained from the Gene Expression Omnibus (GEO) database. R language, along with R packages such as Seurat, was employed to identify cell types based on specific cell markers. Microglia were extracted from the identified cell populations and classified based on their anatomical origin, either gray matter or white matter. Dimensionality reduction and clustering techniques were utilized to identify distinct microglial subpopulations with differential characteristics. Differentially expressed genes (DEGs) between the MS and control groups at the subpopulation level were analyzed by using the Seurat package. Gene set enrichment analysis of Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) was conducted on the DEGs to further explore the biological significance of these differences. Monocle3 was used for pseudotime analysis to study dynamic changes in microglia subpopulations during disease progression. Single cell regulatory network inference and clustering (SCENIC) method was applied to analyze transcription factor (TF) regulatory networks, aiming to identify key transcription factors potentially involved in MS regulation. Results ·After quality control, a total of 149 062 nuclei were retained for analysis. Following dimensional reduction and clustering, 12 238 microglia were identified by using key markers, including DOCK8, CSF1R, P2RY12, and CD74. The results of GO and KEGG pathway analysis showed that in gray matter microglia, functions such as endocytosis, ion homeostasis, and lipid localization were downregulated during disease progression, while in white matter microglia, functions such as protein folding, cytoplasmic translation, and response to thermal stimuli were upregulated. SCENIC analysis revealed that the expression of transcription factors such as FLI1, MITF, and FOXP1 was upregulated in MS. Conclusion ·Microglia play a critical role in MS, with white matter microglia being more significantly impacted by MS than their gray matter counterparts. Transcription factors such as FLI1, MITF, and FOXP1 are identified as key regulators involved in disease modulation, with their associated transcriptional regulatory networks playing a central role in disease modulation.
CAI Qiangwei, SUN Feng, WU Wenyu, SHAO Fuming, GAO Zhengliang, JIN Shengkai. Transcriptional regulatory network analysis of microglia in multiple sclerosis. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2025, 45(1): 29-41 doi:10.3969/j.issn.1674-8115.2025.01.004
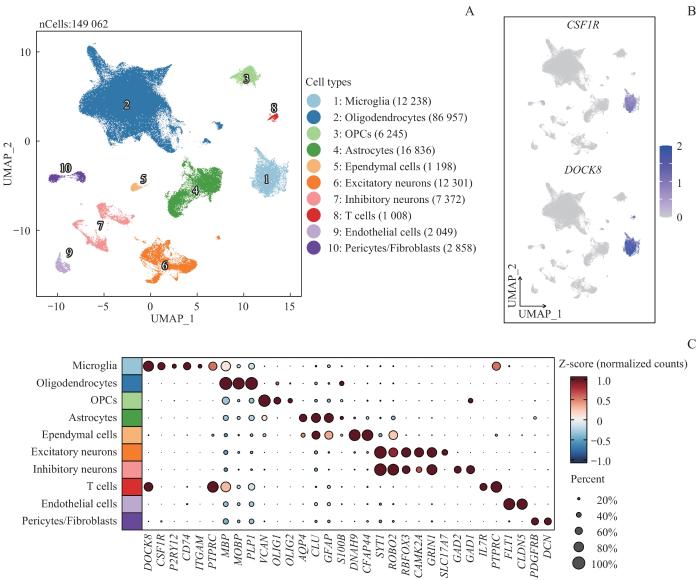
Note: A. UMAP plots of identified cells from MS patients and healthy controls, color-coded by the major cell types. B. Normalized expression of marker genes of microglia. C. Dot plots of marker genes of different cell types.
Fig 1
Dimensionality reduction analysis of snRNA-seq and cell type identification
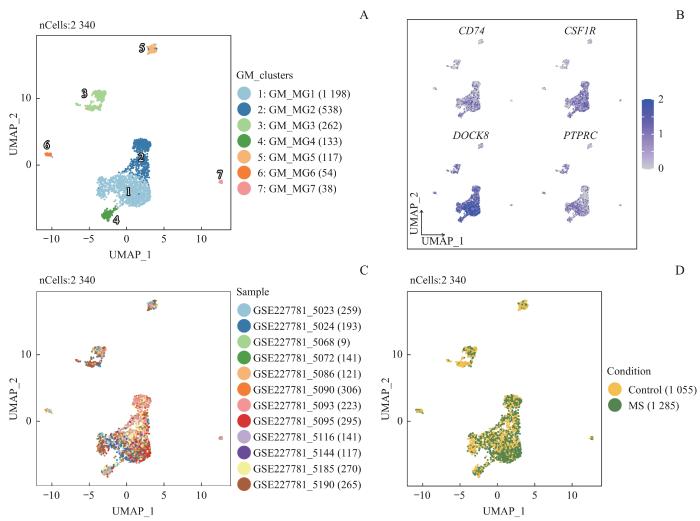
Note: A‒D. UMAP plots of gray matter microglia, color-coded by the subpopulations (A), normalized expression of microglia marker genes (B), samples (C), and MS condition (D).
Fig 2
Heterogeneity analysis of gray matter microglia in MS
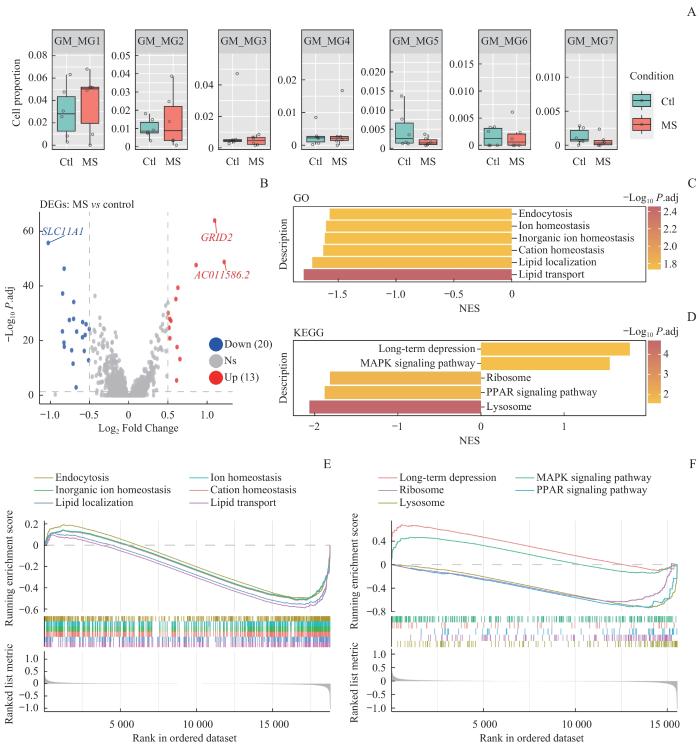
Note: A. Cell proportion of subpopulations of gray matter microglia. Ctl—control. B. Volcano plot of DEGs in gray matter microglia. C/D. Bar plots of GO (C) and KEGG (D) analysis. E/F. Enrichment plots of GO (E) and KEGG (F) analysis.
Fig 3
Subpopulation analysis and enrichment analysis of gray matter microglia in MS
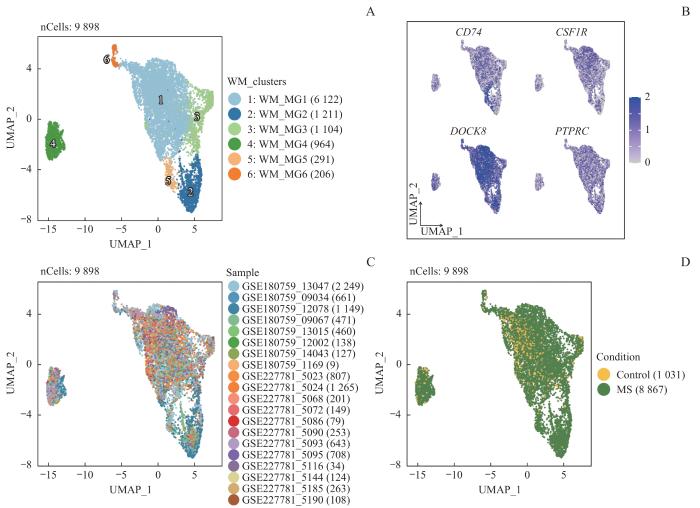
Note: A‒D. UMAP plots of white matter microglia, color-coded by the subpopulations (A), normalized expression of microglia marker genes (B), samples (C), and MS condition (D).
Fig 4
Heterogeneity analysis of white matter microglia in MS
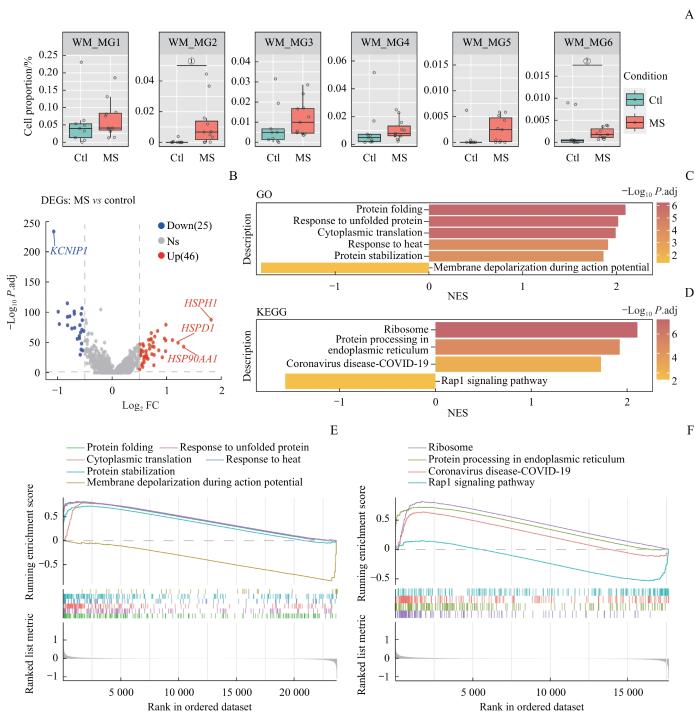
Note: A. Cell proportion of subpopulations of white matter microglia. ①P=0.003, ②P=0.040. B. Volcano plot of DEGs in white matter microglia. C/E. GO analysis of white matter microglia. D/F. KEGG analysis of white matter microglia.
Fig 5
Subpopulation analysis and enrichment analysis of white matter microglia in MS
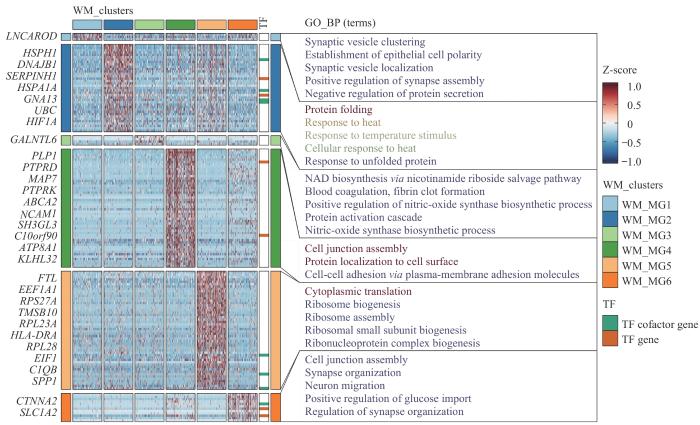
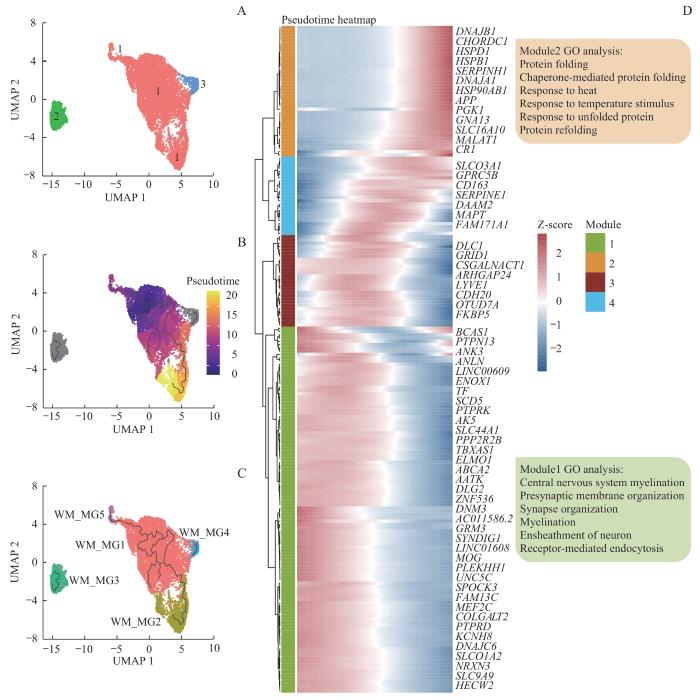
Note: A‒C. UMAP plots of white matter microglia, color-coded by the partition (A), pseudotime (B), and WM_clusters (C). D. Heatmap of gene expression profiles associated with cellular states based on pseudotime analysis. The horizontal axis represents cells arranged according to pseudotime, and the vertical axis displays genes that are correlated with pseudotime.
Fig 7
Pseudotime analysis of white matter microglia
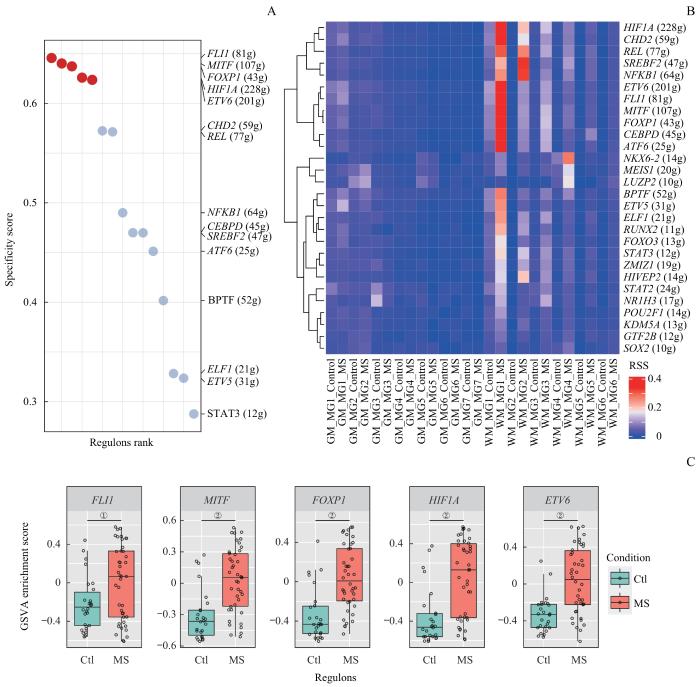
Note: A. Dot plot of RSS ranking among regulons in the MS group. ①P=0.008, ②P=0.000. B. Heatmap of regulon activity identified by SCENIC analysis. C. Box plot of GSVA enrichment analysis score by regulons from GSE179427. g—gene; Ctl—control.
Fig 8
Transcriptional regulatory network analysis of white matter microglia
The study was designed by GAO Zhengliang, JIN Shengkai, CAI Qiangwei, and SUN Feng. Data collection and analysis were conducted by CAI Qiangwei, SUN Feng, WU Wenyu, and SHAO Fuming. The manuscript was drafted by CAI Qiangwei and SUN Feng, and revised by WU Wenyu and SHAO Fuming. Literature review was conducted by WU Wenyu and SHAO Fuming. The research was guided by GAO Zhengliang and JIN Shengkai. All authors have read the final version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclare no relevant conflict of interests.
BENEDICT R H B, AMATO M P, DELUCA J, et al. Cognitive impairment in multiple sclerosis: clinical management, MRI, and therapeutic avenues[J]. Lancet Neurol, 2020, 19(10): 860-871.
KUHLMANN T, MOCCIA M, COETZEE T, et al. Multiple sclerosis progression: time for a new mechanism-driven framework[J]. Lancet Neurol, 2023, 22(1): 78-88.
KENT S A, MIRON V E. Microglia regulation of central nervous system myelin health and regeneration[J]. Nat Rev Immunol, 2024, 24: 49-63.
MCNAMARA N B, MUNRO D A D, BESTARD-CUCHE N, et al. Microglia regulate central nervous system myelin growth and integrity[J]. Nature, 2023, 613: 120-129.
ZHAO S, UMPIERRE A D, WU L J. Tuning neural circuits and behaviors by microglia in the adult brain[J]. Trends Neurosci, 2024, 47(3): 181-194.
SHERAFAT A, PFEIFFER F, REISS A M, et al. Microglial neuropilin-1 promotes oligodendrocyte expansion during development and remyelination by trans-activating platelet-derived growth factor receptor[J]. Nat Commun, 2021, 12: 2265.
DONG Y F, D′MELLO C, PINSKY W, et al. Oxidized phosphatidylcholines found in multiple sclerosis lesions mediate neurodegeneration and are neutralized by microglia[J]. Nat Neurosci, 2021, 24: 489-503.
LAMPRON A, LAROCHELLE A, LAFLAMME N, et al. Inefficient clearance of myelin debris by microglia impairs remyelinating processes[J]. J Exp Med, 2015, 212(4): 481-495.
RAWJI K S, YOUNG A M H, GHOSH T, et al. Niacin-mediated rejuvenation of macrophage/microglia enhances remyelination of the aging central nervous system[J]. Acta Neuropathol, 2020, 139(5): 893-909.
HWANG D, SEYEDSADR M S, ISHIKAWA L L W, et al. CSF-1 maintains pathogenic but not homeostatic myeloid cells in the central nervous system during autoimmune neuroinflammation[J]. Proc Natl Acad Sci USA, 2022, 119(14): e2111804119.
MARZAN D E, BRÜGGER-VERDON V, WEST B L, et al. Activated microglia drive demyelination via CSF1R signaling[J]. Glia, 2021, 69(6): 1583-1604.
TAHMASEBI F, PASBAKHSH P, MORTEZAEE K, et al. Effect of the CSF1R inhibitor PLX3397 on remyelination of corpus callosum in a cuprizone-induced demyelination mouse model[J]. J Cell Biochem, 2019, 120(6): 10576-10586.
VOET S, PRINZ M, VAN LOO G. Microglia in central nervous system inflammation and multiple sclerosis pathology[J]. Trends Mol Med, 2019, 25(2): 112-123.
SEN M K, MAHNS D A, COORSSEN J R, et al. The roles of microglia and astrocytes in phagocytosis and myelination: insights from the cuprizone model of multiple sclerosis[J]. Glia, 2022, 70(7): 1215-1250.
FRISCHER J M, BRAMOW S, DAL-BIANCO A, et al. The relation between inflammation and neurodegeneration in multiple sclerosis brains[J]. Brain, 2009, 132(5): 1175-1189.
CLARK I C, GUTIÉRREZ-VÁZQUEZ C, WHEELER M A, et al. Barcoded viral tracing of single-cell interactions in central nervous system inflammation[J]. Science, 2021, 372(6540): eabf1230.
JÄKEL S, AGIRRE E, MENDANHA FALCÃO A, et al. Altered human oligodendrocyte heterogeneity in multiple sclerosis[J]. Nature, 2019, 566: 543-547.
KISS M G, MINDUR J E, YATES A G, et al. Interleukin-3 coordinates glial-peripheral immune crosstalk to incite multiple sclerosis[J]. Immunity, 2023, 56(7): 1502-1514.e8.
MIEDEMA A, GERRITS E, BROUWER N, et al. Brain macrophages acquire distinct transcriptomes in multiple sclerosis lesions and normal appearing white matter[J]. Acta Neuropathol Commun, 2022, 10(1): 8.
HAFEMEISTER C, SATIJA R. Normalization and variance stabilization of single-cell RNA-seq data using regularized negative binomial regression[J]. Genome Biol, 2019, 20(1): 296.
HU C, LI T, XU Y, et al. CellMarker 2.0: an updated database of manually curated cell markers in human/mouse and web tools based on scRNA-seq data[J]. Nucleic Acids Res, 2023, 51(d1): D870-D876.
TRAPNELL C, CACCHIARELLI D, GRIMSBY J, et al. The dynamics and regulators of cell fate decisions are revealed by pseudotemporal ordering of single cells[J]. Nat Biotechnol, 2014, 32: 381-386.
RITCHIE M E, PHIPSON B, WU D, et al. Limma powers differential expression analyses for RNA-sequencing and microarray studies[J]. Nucleic Acids Res, 2015, 43(7): e47.
KOOI E J, STRIJBIS E M, VAN DER VALK P, et al. Heterogeneity of cortical lesions in multiple sclerosis: clinical and pathologic implications[J]. Neurology, 2012, 79(13): 1369-1376.
FAN L Y, YANG J, LIU R Y, et al. Integrating single-nucleus sequence profiling to reveal the transcriptional dynamics of Alzheimer′s disease, Parkinson's disease, and multiple sclerosis[J]. J Transl Med, 2023, 21(1): 649.
ZINATIZADEH M R, SCHOCK B, CHALBATANI G M, et al. The Nuclear Factor Kappa B (NF-kB) signaling in cancer development and immune diseases[J]. Genes Dis, 2021, 8(3): 287-297.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}