Advances in epigenetic mechanisms of lead toxicity
ZHANG Xinxin1, YAN Chonghuai,1,2
1.Shanghai Jiao Tong University School of Public Health, Shanghai 200025, China
2.Ministry of Education-Shanghai Key Laboratory of Children's Environmental Health, Xinhua Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai 200092, China
铅是一种普遍存在于环境中的有毒重金属,也是人类历史上使用最早、应用最为广泛的重金属元素之一。由于铅在环境中不可降解,并且在人体内具有较长的生物累积效应(可长达30~50年),即使极低浓度的铅也能对人体造成健康损害,因而被世界卫生组织(World Health Organization,WHO)列为十大公共卫生关注化学品之一。铅进入人体后,通常会分布在脑、肝脏、肾脏、牙齿和骨骼等组织中,进而对全身各个系统、多种脏器和组织产生广泛的毒性作用。表观遗传学是研究基因表达在不改变核苷酸序列的情况下发生可遗传变化的学科,它揭示了基因表达修饰如何对细胞进行调控,导致具有相同DNA序列的细胞表现出不同形态与功能。尽管铅的毒性机制尚未完全明确,但近年来的研究表明,表观遗传学调控可能是铅毒性作用的重要机制之一。环境铅暴露可通过引发个体细胞的DNA甲基化、组蛋白修饰和微RNA(microRNA,miRNA)等表观遗传学改变,进而而诱发多种毒性反应。该文就铅毒性相关的表观遗传学机制研究现状,着重从DNA甲基化、组蛋白修饰和miRNA 3个方面进行综述,旨在从表观遗传学角度审视铅毒性,并为进一步探究铅的毒性机制提供理论基础。
关键词:铅
;
DNA甲基化
;
组蛋白修饰
;
微RNA
Abstract
Lead is a ubiquitous toxic heavy metal and one of the earliest and most widely used heavy metal elements in human history. Due to its non-degradable nature in the environment and its long biological accumulation effects (lasting up to 30‒50 years) in the human body, even trace amounts of lead can cause significant health damage. It has therefore been classified as one of the top ten public health concerns by the World Health Organization (WHO). Once absorbed into the body, lead is typically distributed in tissues such as the brain, liver, kidneys, teeth, and bones, thereby exerting widespread toxic effects on multiple organ systems. Epigenetics is the study of heritable changes in gene expression that occur without alterations in the nucleotide sequence. It reveals how modifications in gene expression regulate cellular functions, leading to diverse cellular phenotypes and functions despite identical DNA sequences. Although the toxic mechanisms of lead are not yet fully elucidated, recent studies suggest that epigenetic regulation may play a significant role in mediating lead toxicity. Environmental lead exposure can induce various epigenetic modifications in cells, such as DNA methylation, histone modifications, and microRNA (miRNA) alterations, which, in turn, can trigger multiple toxic responses. This paper presents a concise overview of current epigenetic investigations into lead toxicity, emphasizing DNA methylation, histone modifications, and miRNA dynamics. By adopting an epigenetic perspective, it offers a theoretical framework into understanding lead's toxic mechanisms comprehensively, facilitating further research in prevention and treatment strategies.
ZHANG Xinxin, YAN Chonghuai. Advances in epigenetic mechanisms of lead toxicity. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2025, 45(4): 500-507 doi:10.3969/j.issn.1674-8115.2025.04.013
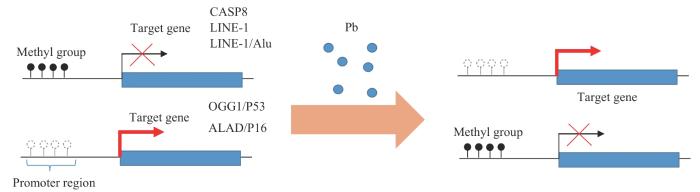
Fig 1
Dual effects of lead on DNA methylation and downstream genes
长散在重复序列1(long interspersed nuclear elements 1,LINE1)和Alu重复序列承载了人类基因组中大部分的CpG位点,其甲基化水平常被用以替代评估全基因组甲基化状况[17]。铅暴露通常导致重复序列的整体低甲基化。在中国职业铅暴露的相关研究[18-19]中,职业工人的血铅水平与LINE1及Alu的甲基化水平显著负相关。尽管仍很难确定铅暴露导致的DNA甲基化水平下降是疾病的原因还是结果,但已有研究[1]表明,这种低甲基化是一些复杂疾病的特征,它与染色质松弛和转录激活相关,导致基因组不稳定,进而增加癌症风险。
铅暴露也会导致特定位点基因启动子的高甲基化。在以12.5 mg/kg(每周3次)醋酸铅灌胃处理的10周龄Sprague-Dawley大鼠大脑皮质中,有研究观察到抑癌基因P53和DNA修复蛋白8-氧鸟嘌呤DNA糖苷酶(8-oxoguanine DNA glycosylase,OGG1)基因启动子的高甲基化[20];前者具有控制细胞周期进程、保护DNA完整性、预防肿瘤发生的作用,后者参与DNA碱基修复,表达调控相关基因的表达,二者基因启动子的高甲基化可能与实验中铅暴露鼠大脑皮层细胞凋亡增加及DNA损伤加剧等现象有关。这一现象也同样在职业暴露工人的人群研究中得以复现[19]。此外,铅暴露也与δ-氨基-γ-酮戊酸脱氢酶(δ-aminolevulinic acid dehydratase,ALAD)[21]以及P16[18]基因启动子高甲基化相关。ALAD的抑制可导致δ-氨基-γ-酮戊酸(δ-aminolevulinic acid,ALA)蓄积,进而引发血液系统的氧化损伤;而P16基因在细胞周期调控中发挥重要作用,其表达抑制可能增加肿瘤发生的风险。总之,铅引起的特定位点高甲基化主要与抑癌基因表达抑制和氧化应激损伤等效应密切相关。
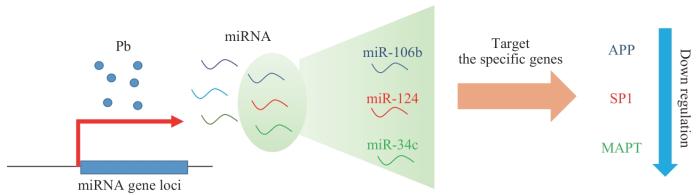
铅暴露常伴随miRNA的表达改变。有研究发现墨西哥女性血铅与miR-155表达呈显著正相关,与miR-126表达呈显著负相关[46],WEN等人[47]结合欧盟Interreg Memories项目中5例阿尔茨海默病(Alzheimer's disease,AD)患者和2例对照组脑区miRNA数据及瑞典北部健康与疾病研究(Northern Sweden Health and Disease Study,NSHDS)受试者健康时的血液miRNA测序结果,发现铅暴露与miR-3651、miR-150-5p以及miR-664b-3p表达呈负相关,与miR-627表达呈正相关。在中国铅暴露职业工人样本中,miR-572表达与血铅水平呈显著正相关,miR-520c-3p、miR-211和miR-148a的表达与血铅水平呈显著负相关[45]。
Fig 2
Lead Exposure alters AD-related miRNAs and suppresses gene expression
铅的神经系统毒性还可通过其伴随的miRNA表达变化及其诱导神经突触结构与功能改变来解释。有研究发现,高血铅诱导大鼠海马miR-34b表达增高以及突触相关膜蛋白2(vesicle-associated membrane protein 2,VAMP2)的下调,导致发育性神经精神功能障碍,阻断miR-34b可减轻铅对VAMP2的影响[57]。在另一项研究中,WANG等[58]发现,阻断miR-130可逆转铅诱导的前扣带回皮层突触前靶标SNAP-25蛋白表达降低,并减轻铅暴露后的突触前结构与功能改变以及小鼠注意力缺陷。目前已有的铅暴露对miRNA的调控及其可能的功能总结详见表1。
Tab 1
表1
表1铅暴露对miRNA的调控及其相关效应
Tab 1 Regulation of miRNA by Pb exposure and its related effects
Gene
Expression change
Sample type
Effect
Reference
miR-155/miR-221
Upregulated
Peripheral blood samples from adults
Cell apoptosis, cell proliferation, and cytokine production
PMID: 32902755
miR-146a
Upregulated
Peripheral blood samples from adults
Inhibition of inflammatory factor release and cell apoptosis
PMID: 36274319
miR-106b-56p
Upregulated
HT-22 and PC12 cell lines
Decreased XIAP levels and cell viability
PMID: 32344020
miR-378a-3p
Upregulated
HT-22 cell line
Reduced GSH and increased lipid ROS levels
PMID: 35588615
miR-143-5p
Downregulated
Fibroblast samples
Regulation of dysfunction of interstitial fibroblasts
PMID: 35485286
miR-106b/miR-124/miR-34c
Upregulated
Cerebral cortex tissue samples
Induced neurotoxicity and learning/memory impairment
ZHANG Xinxin was responsible for literature review, drafting the initial manuscript, and completing revisions, while YAN Chonghuai was responsible for providing writing guidance and reviewing the paper. Both authors have read and agreed to submit the final manuscript.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
Both authors disclose no relevant conflict of interests.
APPLETON A A, JACKSON B P, KARAGAS M, et al. Prenatal exposure to neurotoxic metals is associated with increased placental glucocorticoid receptor DNA methylation[J]. Epigenetics, 2017, 12(8): 607-615.
OKAMOTO Y, IWAI-SHIMADA M, NAKAI K, et al. Global DNA methylation in cord blood as a biomarker for prenatal lead and antimony exposures[J]. Toxics, 2022, 10(4): 157.
ZENG Z J, HUO X, ZHANG Y, et al. Differential DNA methylation in newborns with maternal exposure to heavy metals from an e-waste recycling area[J]. Environ Res, 2019, 171: 536-545.
BOZACK A K, RIFAS-SHIMAN S L, COULL B A, et al. Prenatal metal exposure, cord blood DNA methylation and persistence in childhood: an epigenome-wide association study of 12 metals[J]. Clin Epigenetics, 2021, 13(1): 208.
TUNG P W, KENNEDY E M, BURT A, et al. Prenatal lead (Pb) exposure is associated with differential placental DNA methylation and hydroxymethylation in a human population[J]. Epigenetics, 2022, 17(13): 2404-2420.
RYGIEL C A, DOLINOY D C, PERNG W, et al. Trimester-specific associations of prenatal lead exposure with infant cord blood DNA methylation at birth[J]. Epigenet Insights, 2020, 13: 2516865720938669.
RYGIEL C A, DOLINOY D C, BAKULSKI K M, et al. DNA methylation at birth potentially mediates the association between prenatal lead (Pb) exposure and infant neurodevelopmental outcomes[J]. Environ Epigenet, 2021, 7(1): dvab005.
WANG K, LIU S Y, SVOBODA L K, et al. Tissue- and sex-specific DNA methylation changes in mice perinatally exposed to lead (Pb)[J]. Front Genet, 2020, 11: 840.
SVOBODA L K, NEIER K R, WANG K, et al. Tissue and sex-specific programming of DNA methylation by perinatal lead exposure: implications for environmental epigenetics studies[J]. Epigenetics, 2021, 16(10): 1102-1122.
SVOBODA L K, WANG K, JONES T R, et al. Sex-specific alterations in cardiac DNA methylation in adult mice by perinatal lead exposure[J]. Int J Environ Res Public Health, 2021, 18(2): 577.
SVOBODA L K, WANG K, GOODRICH J M, et al. Perinatal lead exposure promotes sex-specific epigenetic programming of disease-relevant pathways in mouse heart[J]. Toxics, 2023, 11(1): 85.
MORGAN R K, WANG K, SVOBODA L K, et al. Effects of developmental lead and phthalate exposures on DNA methylation in adult mouse blood, brain, and liver identifies tissue- and sex-specific changes with implications for genomic imprinting[J]. bioRxiv, 2023: 2023.09.29.560131.
MCCABE C, ANDERSON O S, MONTROSE L, et al. Sexually dimorphic effects of early-life exposures to endocrine disruptors: sex-specific epigenetic reprogramming as a potential mechanism[J]. Curr Environ Health Rep, 2017, 4(4): 426-438.
WANG K, MENG Y, WANG T W, et al. Global and gene-specific promoter methylation, and micronuclei induction in lead-exposed workers: a cross-sectional study[J]. Environ Mol Mutagen, 2021, 62(7): 428-434.
WANG T W, MENG Y, TU Y T, et al. Associations between DNA methylation and genotoxicity among lead-exposed workers in China[J]. Environ Pollut, 2023, 316(Pt 1): 120528.
EL-SHETRY E S, MOHAMED A A, KHATER S I, et al. Synergistically enhanced apoptotic and oxidative DNA damaging pathways in the rat brain with lead and/or aluminum metals toxicity: expression pattern of genes OGG1 and P53[J]. J Trace Elem Med Biol, 2021, 68: 126860.
YOHANNES Y B, NAKAYAMA S M, YABE J, et al. Blood lead levels and aberrant DNA methylation of the ALAD and p16 gene promoters in children exposed to environmental-lead[J]. Environ Res, 2020, 188: 109759.
COLICINO E, JUST A, KIOUMOURTZOGLOU M A, et al. Blood DNA methylation biomarkers of cumulative lead exposure in adults[J]. J Expo Sci Environ Epidemiol, 2021, 31(1): 108-116.
PAUL K C, HORVATH S, DEL ROSARIO I, et al. DNA methylation biomarker for cumulative lead exposure is associated with Parkinson's disease[J]. Clin Epigenetics, 2021, 13(1): 59.
LIEBERMAN-CRIBBIN W, DOMINGO-RELLOSO A, NAVAS-ACIEN A, et al. Epigenetic biomarkers of lead exposure and cardiovascular disease: prospective evidence in the strong heart study[J]. J Am Heart Assoc, 2022, 11(23): e026934.
SÁNCHEZ O F, LIN L F, XIE J K, et al. Lead exposure induces dysregulation of constitutive heterochromatin hallmarks in live cells[J]. Curr Res Toxicol, 2022, 3: 100061.
GU X Z, XU Y, XUE W Z, et al. Interplay of miR-137 and EZH2 contributes to the genome-wide redistribution of H3K27me3 underlying the Pb-induced memory impairment[J]. Cell Death Dis, 2019, 10(9): 671.
XIAO J, WANG T, XU Y, et al. Long-term probiotic intervention mitigates memory dysfunction through a novel H3K27me3-based mechanism in lead-exposed rats[J]. Transl Psychiatry, 2020, 10(1): 25.
EID A, BIHAQI S W, RENEHAN W E, et al. Developmental lead exposure and lifespan alterations in epigenetic regulators and their correspondence to biomarkers of Alzheimer's disease[J]. Alzheimers Dement (Amst), 2016, 2: 123-131.
VARMA G, SOBOLEWSKI M, CORY-SLECHTA D A, et al. Sex- and brain region- specific effects of prenatal stress and lead exposure on permissive and repressive post-translational histone modifications from embryonic development through adulthood[J]. Neurotoxicology, 2017, 62: 207-217.
LIN L F, XIE J K, SÁNCHEZ O F, et al. Low dose lead exposure induces alterations on heterochromatin hallmarks persisting through SH-SY5Y cell differentiation[J]. Chemosphere, 2021, 264(Pt 1): 128486.
LUO M, XU Y, CAI R, et al. Epigenetic histone modification regulates developmental lead exposure induced hyperactivity in rats[J]. Toxicol Lett, 2014, 225(1): 78-85.
XU L H, MU F F, ZHAO J H, et al. Lead induces apoptosis and histone hyperacetylation in rat cardiovascular tissues[J]. PLoS One, 2015, 10(6): e0129091.
KIRAN G S, KUMAR P K, MITRA P, et al. Unravelling blood-based epigenetic mechanisms: the impact of hsa-miR-146a and histone H3 acetylation in lead-induced inflammation among occupational workers[J]. Int Arch Occup Environ Health, 2023, 96(9): 1257-1266.
WANG Y W, HU Y Z, WU Z T, et al. Latent role of in vitro Pb exposure in blocking Aβ clearance and triggering epigenetic modifications[J]. Environ Toxicol Pharmacol, 2019, 66: 14-23.
CHESHMAZAR N, HAMZEH-MIVEHROUD M, NOZAD CHAROUDEH H, et al. Current trends in development of HDAC-based chemotherapeutics[J]. Life Sci, 2022, 308: 120946.
ZHOU R Q, ZHAO J, LI D Y, et al. Combined exposure of lead and cadmium leads to the aggravated neurotoxicity through regulating the expression of histone deacetylase 2[J]. Chemosphere, 2020, 252: 126589.
GU X Z, HUANG X Y, LI D Y, et al. Nuclear accumulation of histone deacetylase 4 (HDAC4) by PP1-mediated dephosphorylation exerts neurotoxicity in Pb-exposed neural cells[J]. Neurotoxicology, 2020, 81: 395-405.
XU M, YU Z M, HU F F, et al. Identification of differential plasma miRNA profiles in Chinese workers with occupational lead exposure[J]. Biosci Rep, 2017, 37(5): BSR20171111.
OCHOA-MARTÍNEZ Á C, VARELA-SILVA J A, ORTA-GARCÍA S T, et al. Lead (Pb) exposure is associated with changes in the expression levels of circulating miRNAS (miR-155, miR-126) in Mexican women[J]. Environ Toxicol Pharmacol, 2021, 83: 103598.
WEN Q F, VERHEIJEN M, WITTENS M M J, et al. Lead-exposure associated miRNAs in humans and Alzheimer's disease as potential biomarkers of the disease and disease processes[J]. Sci Rep, 2022, 12(1): 15966.
MITRA P, GOYAL T, SINGH P, et al. Assessment of circulating miR-20b, miR-221, and miR-155 in occupationally lead-exposed workers of North-Western India[J]. Environ Sci Pollut Res Int, 2021, 28(3): 3172-3181.
XUE C, KANG B P, SU P, et al. microRNA-106b-5p participates in lead (Pb2+)-induced cell viability inhibition by targeting XIAP in HT-22 and PC12 cells[J]. Toxicol In Vitro, 2020, 66: 104876.
WANG W X, SHI F, CUI J M, et al. miR-378a-3p/SLC7A11 regulate ferroptosis in nerve injury induced by lead exposure[J]. Ecotoxicol Environ Saf, 2022, 239: 113639.
HAN L, ZOU Y F, YU C. Targeting CC chemokine ligand (CCL) 20 by miR-143-5p alleviate lead poisoning-induced renal fibrosis by regulating interstitial fibroblasts excessive proliferation and dysfunction[J]. Bioengineered, 2022, 13(4): 11156-11168.
MASOUD A M, BIHAQI S W, ALANSI B, et al. Altered microRNA, mRNA, and protein expression of neurodegeneration-related biomarkers and their transcriptional and epigenetic modifiers in a human tau transgenic mouse model in response to developmental lead exposure[J]. J Alzheimers Dis, 2018, 63(1): 273-282.
DASH M, EID A, SUBAIEA G, et al. Developmental exposure to lead (Pb) alters the expression of the human tau gene and its products in a transgenic animal model[J]. NeuroToxicology, 2016, 55: 154-159.
WANG R K, WU Z T, LIU R D, et al. Age-related miRNAs dysregulation and abnormal BACE1 expression following Pb exposure in adolescent mice[J]. Environ Toxicol, 2022, 37(8): 1902-1913.
LIU R D, WANG Y W, BAI L, et al. Time-course miRNA alterations and SIRT1 inhibition triggered by adolescent lead exposure in mice[J]. Toxicol Res (Camb), 2021, 10(4): 667-676.
YANG C H, KANG B P, CAO Z P, et al. Early-life Pb exposure might exert synapse-toxic effects via inhibiting synapse-associated membrane protein 2 (VAMP2) mediated by upregulation of miR-34b[J]. J Alzheimers Dis, 2022, 87(2): 619-633.
WANG T, GUAN R L, ZOU Y F, et al. miR-130/SNAP-25 axis regulate presynaptic alteration in anterior cingulate cortex involved in lead induced attention deficits[J]. J Hazard Mater, 2023, 443(Pt B): 130249.
... 铅暴露常伴随miRNA的表达改变.有研究发现墨西哥女性血铅与miR-155表达呈显著正相关,与miR-126表达呈显著负相关[46],WEN等人[47]结合欧盟Interreg Memories项目中5例阿尔茨海默病(Alzheimer's disease,AD)患者和2例对照组脑区miRNA数据及瑞典北部健康与疾病研究(Northern Sweden Health and Disease Study,NSHDS)受试者健康时的血液miRNA测序结果,发现铅暴露与miR-3651、miR-150-5p以及miR-664b-3p表达呈负相关,与miR-627表达呈正相关.在中国铅暴露职业工人样本中,miR-572表达与血铅水平呈显著正相关,miR-520c-3p、miR-211和miR-148a的表达与血铅水平呈显著负相关[45]. ...
1
... 铅暴露常伴随miRNA的表达改变.有研究发现墨西哥女性血铅与miR-155表达呈显著正相关,与miR-126表达呈显著负相关[46],WEN等人[47]结合欧盟Interreg Memories项目中5例阿尔茨海默病(Alzheimer's disease,AD)患者和2例对照组脑区miRNA数据及瑞典北部健康与疾病研究(Northern Sweden Health and Disease Study,NSHDS)受试者健康时的血液miRNA测序结果,发现铅暴露与miR-3651、miR-150-5p以及miR-664b-3p表达呈负相关,与miR-627表达呈正相关.在中国铅暴露职业工人样本中,miR-572表达与血铅水平呈显著正相关,miR-520c-3p、miR-211和miR-148a的表达与血铅水平呈显著负相关[45]. ...
1
... 铅暴露常伴随miRNA的表达改变.有研究发现墨西哥女性血铅与miR-155表达呈显著正相关,与miR-126表达呈显著负相关[46],WEN等人[47]结合欧盟Interreg Memories项目中5例阿尔茨海默病(Alzheimer's disease,AD)患者和2例对照组脑区miRNA数据及瑞典北部健康与疾病研究(Northern Sweden Health and Disease Study,NSHDS)受试者健康时的血液miRNA测序结果,发现铅暴露与miR-3651、miR-150-5p以及miR-664b-3p表达呈负相关,与miR-627表达呈正相关.在中国铅暴露职业工人样本中,miR-572表达与血铅水平呈显著正相关,miR-520c-3p、miR-211和miR-148a的表达与血铅水平呈显著负相关[45]. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}