Analysis of m6A methylation expression profiles in liver tissue of high-fat diet-induced mouse models of NAFLD
LIU Junjun,1,2, LU Sumei2, ZHANG Bingyang2, LI Yongqing2, MA Wanshan,1,2
1.School of Clinical Medicine, Shandong University, Jinan 250014, China
2.Department of Clinical Laboratory Medicine, The First Affiliated Hospital of Shandong First Medical University/Shandong Provincial Qianfoshan Hospital, Shandong Medicine and Health Key Laboratory of Laboratory Medicine, Jinan 250014, China
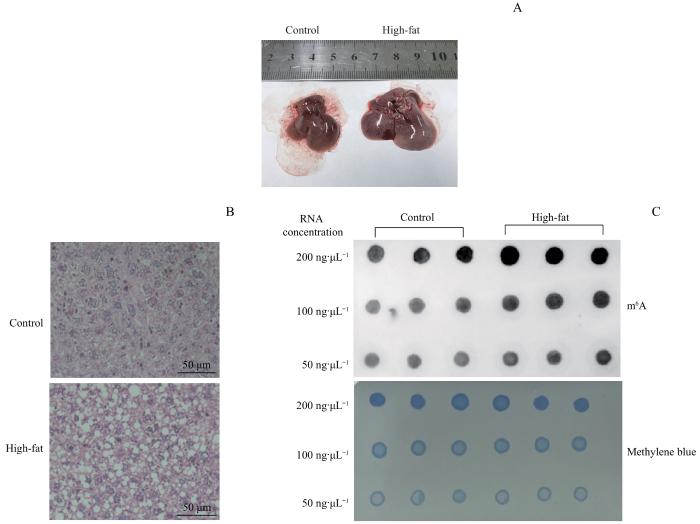
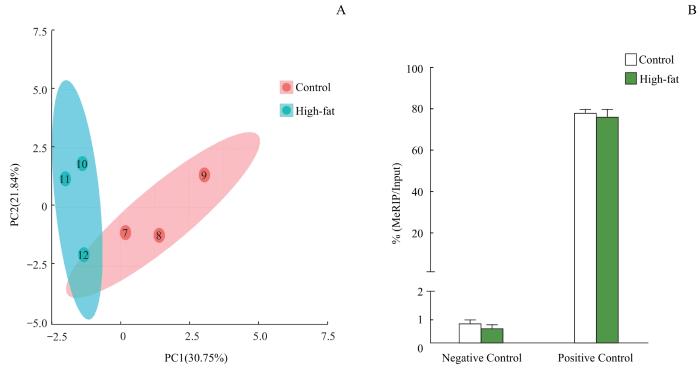
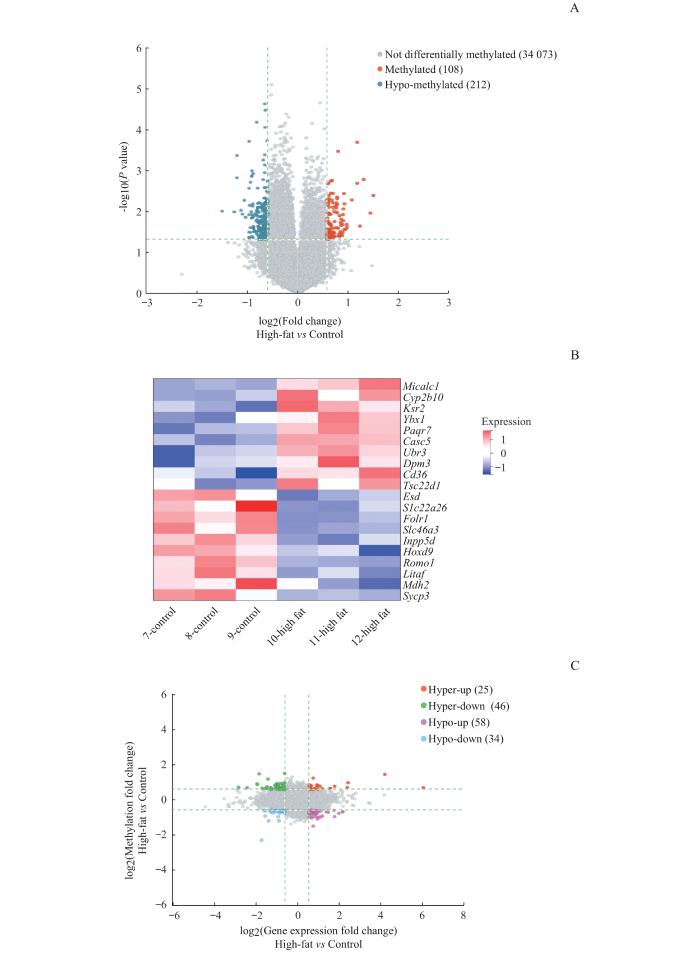
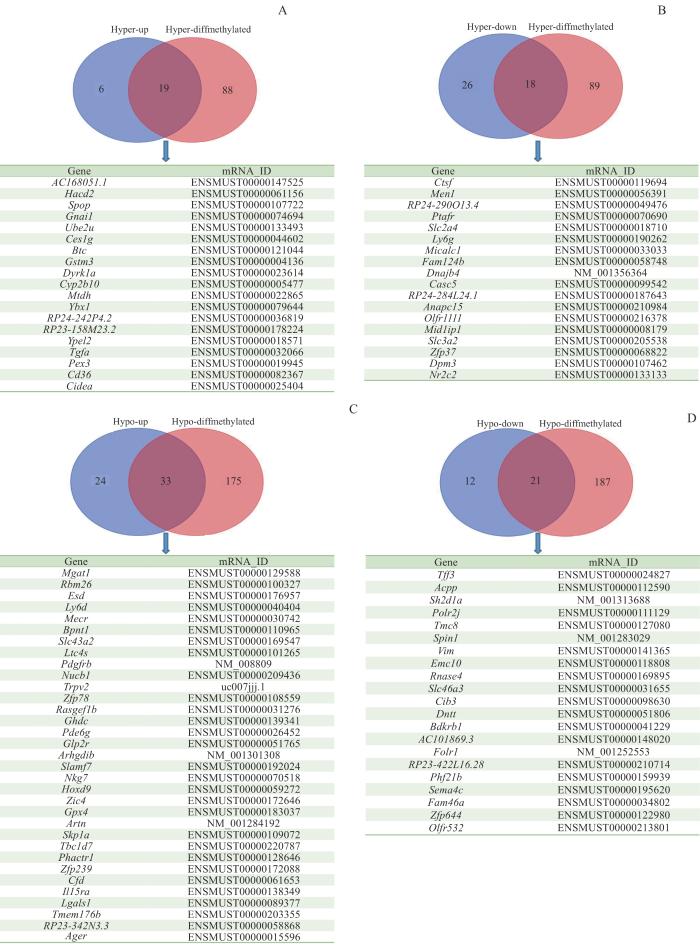
Objective ·To detect the differences in m6A methylation modification and gene expression of liver tissue mRNA in high-fat diet-induced mouse models of non-alcoholic fatty liver disease (NAFLD) using microarray technology. Methods ·The NAFLD models were established in 6-8 weeks old male C57BL/6J mice fed with high-fat chow for 16 weeks (high-fat group, n=10). The basal group (n=10) was given 10% fat diet. Hematoxylin-eosin (H-E) staining was used to assess the histopathological changes in liver tissue and to determine the success of the NAFLD models. The changes of mRNA m6A methylation and expression levels in the liver tissues of the two groups were detected by using methylated RNA immunoprecipitation (MeRIP) and microarray expression profiling. Results ·The livers of the mice in the basal group were bright red with few fat deposits, while the livers of the mice in the high-fat group were yellowish with diffuse infiltration and fusion of lipid droplets in the hepatocytes by H-E staining, suggesting that the high-fat diet-induced NAFLD models were successfully constructed. The results of the MeRIP-microarray showed that the m6A methylation levels of 320 genes in the livers of mice in the high-fat group were significantly altered compared with those in the basal group (P<0.05 and fold change>1.5), of which 108 genes were up-regulated and 212 genes were down-regulated. Genes with significant differences in m6A methylation levels between the two groups were intersected with those with differentially expressed mRNAs, and 163 genes were found to have significant differences in both m6A methylation level and mRNA expression level. Conclusion ·The change in m6A modification of liver tissue mRNA in the high-fat diet-induced mouse models of NAFLD is significant and the change is associated with the gene expression of mRNA.
LIU Junjun, LU Sumei, ZHANG Bingyang, LI Yongqing, MA Wanshan. Analysis of m6A methylation expression profiles in liver tissue of high-fat diet-induced mouse models of NAFLD. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(10): 1227-1235 doi:10.3969/j.issn.1674-8115.2023.10.002
Note: A. General observation of the livers. B.H-E staining showed the cytoplasm of liver tissue in the high-fat group was filled with lipid droplets. C.Dot-blot showed the increase of m6A methylation levels in the high-fat group.
Fig 1
Changes of m6A modification levels in high-fat diet mouse livers
Note: A. Volcano plot of differential m6A methylation mRNAs. B.Cluster diagram showed top 10 differentially m6A-methylated mRNAs. C.Four quadrant diagrams showed differential methylation associated with gene expression in the high-fat group and the control group.
Fig 3
Conjoint analysis of differential m6A methylation mRNAs and differentially expressed genes in the livers of mice
The study was designed by MA Wanshan, LU Sumei and LIU Junjun. The manuscript was drafted and revised by MA Wanshan, LU Sumei and LIU Junjun. The experiments were performed by LIU Junjun, ZHANG Bingyang and LI Yongqing. The data were analyzed by LIU Junjun, ZHANG Bingyang and LI Yongqing. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
All authors disclose no relevant conflict of interests.
XIANG W. IL-17A deficiency alleviates LPS-induced steatotic liver injury in mice and the underlying mechanism[D]. Chongqing: Third Military Medical University of Chinese P.L.A., 2017.
ESTES C, ANSTEE Q M, ARIAS-LOSTE M T, et al. Modeling NAFLD disease burden in China, France, Germany, Italy, Japan, Spain, United Kingdom, and United States for the period 2016‒2030[J]. J Hepatol, 2018, 69(4): 896-904.
CHENG Y, HOU T, PING J, et al. Quantitative succinylome analysis in the liver of non-alcoholic fatty liver disease rat model[J]. Proteome Sci, 2016, 14: 3.
XU Z J, SHI J P, YU D R, et al. Evaluating the relationship between metabolic syndrome and liver biopsy-proven non-alcoholic steatohepatitis in China: a multicenter cross-sectional study design[J]. Adv Ther, 2016, 33(11): 2069-2081.
YOUNOSSI Z M, KOENIG A B, ABDELATIF D, et al. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes[J]. Hepatology, 2016, 64(1): 73-84.
YOUNOSSI Z M, GOLABI P, DE AVILA L, et al. The global epidemiology of NAFLD and NASH in patients with type 2 diabetes: a systematic review and meta-analysis[J]. J Hepatol, 2019, 71(4): 793-801.
PASCHOS P, PALETAS K. Non alcoholic fatty liver disease and metabolic syndrome[J]. Hippokratia, 2009, 13(1): 9-19.
STEFAN N, HÄRING H U, CUSI K. Non-alcoholic fatty liver disease: causes, diagnosis, cardiometabolic consequences, and treatment strategies[J]. Lancet Diabetes Endocrinol, 2019, 7(4): 313-324.
ALEXANDER M, LOOMIS A K, VAN DER LEI J, et al. Risks and clinical predictors of cirrhosis and hepatocellular carcinoma diagnoses in adults with diagnosed NAFLD: real-world study of 18 million patients in four European cohorts[J]. BMC Med, 2019, 17(1): 95.
WANG Y Y, JIANG J P. Research progress of the relationship between nonalcoholic fatty liver disease and components of metabolic syndrome[J]. China Modern Medicine, 2022, 29(15): 36-39, 43.
LI Y Q. Role and mechanism of METTL3-mediated CYP2B6 m6A methylation modification in insulin sensitivity in hepatocytes with non-alcoholic fatty liver disease[D]. Jinan: Shandong University, 2022.
YE D W, MA W S, LU S M. Advances in the study of RNA m6A methylation modification in the metabolic syndrome[J].Chinese Journal of Clinical Laboratory Science, 2022,40(9): 691-694.
CHEN Z Y, PU R, DENG S, et al. N6-methyladenosine methylation and its regulation in metabolic diseases[J]. Chinese Journal of Tissue Engineering Research, 2022, 26(20):3250-3255.
TONG X W, YE X Y, LIU C Y, et al. Relationship between mRNA expression of PPARG, CIDEA, ECHDC3, CGN and lipid levels in peripheral blood of patients with type 2 diabetes mellitus complicated with obesity[J].Shenzhen Journal of Integrated Traditional Chinese and Western Medicine, 2021, 31(6): 4-8, 199.
SANS A, BONNAFOUS S, ROUSSEAU D, et al. The differential expression of CIDE family members is associated with NAFLD progression from steatosis to steatohepatitis[J]. Sci Rep, 2019, 9(1): 7501.
QUIROGA A D, LI L, TRÖTZMÜLLER M, et al. Deficiency of carboxylesterase 1/esterase-x results in obesity, hepatic steatosis, and hyperlipidemia[J]. Hepatology, 2012, 56(6): 2188-2198.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}