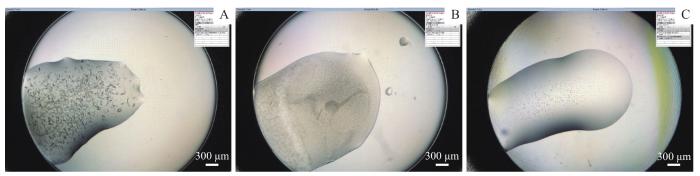
Objective ·To prepare a high-purity cyanobacterial serine protease inhibitor, screen its target proteases, and detect its inhibitory activity. Methods ·A novel serine protease inhibitor from Arthrospira platensis was identified in the Expanded Human Oral Microbiome Database (eHOMD) by amino acid sequence alignment and named as arthropin. The fusion expression vector pSUMO3-arthropin was constructed and transferred into Escherichia coli (E. coli) BL21(DE3) system for fusion protein expression. The recombinant arthropin was purified by a four-step chromatographic purification approach of nickel affinity chromatography, enzymatic digestion, reverse nickel affinity chromatography, and anion exchange chromatography. In addition, the recombinant arthropin was co-incubated with 14 serine proteases such as activated factor Ⅸ (FⅨa), FⅩa, FⅪa, activated protein C (APC) and kallikrein 1 (KLK1), respectively, and then analyzed by SDS-PAGE. The inhibitory rate of arthropin on KLK1 was assayed with kinetic methods. The crystallization conditions of the recombinant arthropin were screened preliminarily, and the suitable crystals were picked for X-ray diffraction to collect the data. Finally, a sub-stable structure model of arthropin was predicted with AlphaFlod Colab. Results ·SDS-PAGE showed that the fused arthropin was successfully expressed in the E. coli BL21(DE3) system, and following purification, the high-purity recombinant arthropin, the relative molecular mass of which was similar to the theoretical value (45 800), was obtained. The co-incubation analysis of recombinant arthropin with 14 serine proteases revealed that arthropin was able to form stable covalent complexes with 9 proteases, including FⅩa, APC, FⅨa, FⅪa, trypsin, cathepsin G, KLK1, KLK7 and thrombin. Arthropin inhibited KLK1 with a second-order association rate constant of 1.7×103 L/(mol·s). Moreover, the recombinant arthropin crystalised under the condition of 25% PEG MME 550, 0.1 mol/L MES (pH 6.5) and 0.01 mol/L ZnCl2 , and the crystals preliminarily diffracted to a resolution of 10 Å (1 Å=1×10-10 m). The analysis of the structure predicted by AlphaFlod Colab revealed that arthropin had the classical structural features of the inhibitory serpin. Conclusion ·Arthropin, a serpin from Arthrospira platensis, was successfully obtained with high purity and a broad-spectrum of serine protease inhibition, but at a low inhibitory rate.
XU Jiawei, ZHOU Aiwu, YANG Yufeng. Preparation and target protease identification of a cyanobacterial serine protease inhibitor, arthropin. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(4): 428-436 doi:10.3969/j.issn.1674-8115.2023.04.004
丝氨酸蛋白酶抑制剂(serine protease inhibitor,serpin)是规模最大、分布最广的一类蛋白酶抑制剂超家族,广泛存在于动物、植物、细菌、病毒和古菌中[1-6]。Serpin家族成员多数由330~500个氨基酸残基组成,它们的一级序列同源性不高(约25%),但二级和三级结构极其相似。典型的serpin主要由1个包含8~9个α螺旋(helix A~I)、3个β片层(β-sheet A~C)的保守主体结构和1个暴露在分子表面的反应中心环(reactive center loop,RCL)组成[6]。RCL的长度和序列的多样性决定着serpin靶蛋白酶的特异性。
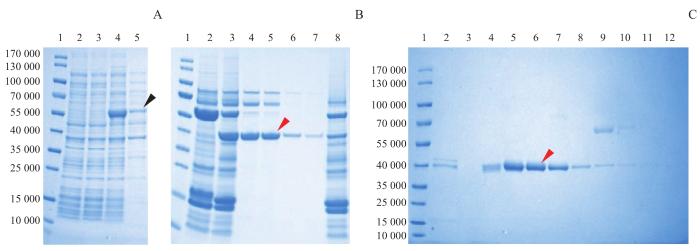
Note:A. Protein expression in the whole bacteria and lysis supernatant of E. coli before and after induction. Lane 1—marker; lane 2—total bacterial protein sample before induction; lane 3—supernatant protein sample before induction; lane 4—total bacterial protein sample after induction for 12 h; lane 5—supernatant protein sample after induction for 12 h. B. Enzymatic digestion and purification by nickel ion affinity chromatography. Lane 1—marker; lane 2—protein sample before SENP2 digestion; lane 3—protein sample after SENP2 digestion; lane 4‒5—flow-through solution of the nickel column after SENP2 digestion; lane 6‒7—washing solution of the nickel column after SENP2 digestion; lane 8—elution solution of the nickel column after SENP2 digestion. C. Purification by anion-exchange chromatography. Lane 1—marker; lane 2‒12—protein samples eluted by a continuous gradient of 145‒685 mmol/L NaCl. The black arrow indicates the fusion protein and the red arrows indicate the monomeric protein.
Fig 2
Expression, enzymatic digestion and purification of arthropin fusion protein analyzed by SDS-PAGE
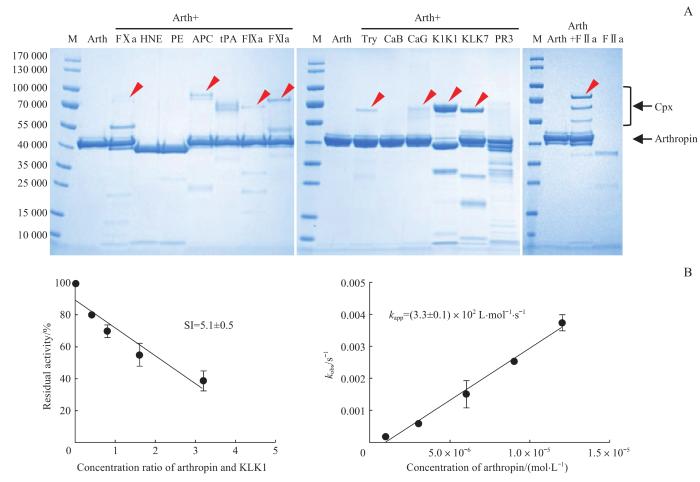
Note:A. Arthropin (3 μg) was incubated with the proteases (0.4 μg), respectively, and the complex formation was analyzed by SDS-PAGE. The red arrows indicate the complexes of arthropin with the target proteases. M—marker; Try—trypsin; Arth—arthropin; Cpx—arthropin-proteinase covalent complex. B. Determination of the SI (left panel) and the kapp (right panel) of arthropin's inhibition of KLK1.
Fig 3
Screening and reaction kinetics assays for target proteases of arthropin
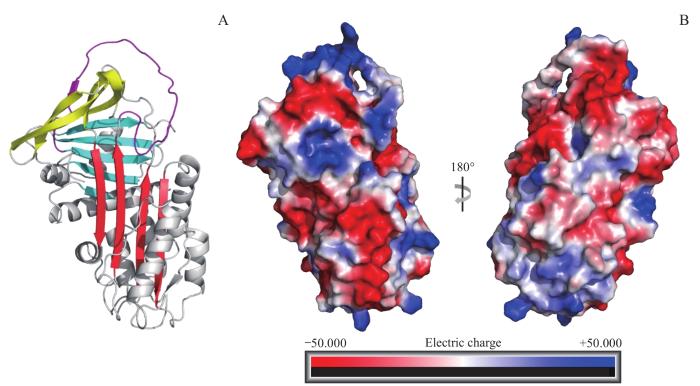
Note:A. The arthropin model created with AlphaFlod Colab. β-sheets A, B, and C are indicated in red, blue, and yellow, respectively. RCL is in purple. B. Surface charge diagram of arthropin. The red colour indicates the negative charge and the blue colour indicates the positive charge.
ZHOU Aiwu conducted and designed this study. XU Jiawei completed all the experiments and the data compilation with the guidance of ZHOU Aiwu and YANG Yufeng. XU Jiawei wrote and revised the thesis, and YANG Yufeng and ZHOU Aiwu provided guidance on the revision of the thesis. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
HUBER R, CARRELL R W. Implications of the three-dimensional structure of α1-antitrypsin for structure and function of serpins[J]. Biochemistry, 1989, 28(23): 8951-8966.
SPENCE M A, MORTIMER M D, BUCKLE A M, et al. A comprehensive phylogenetic analysis of the serpin superfamily[J]. Mol Biol Evol, 2021, 38(7): 2915-2929.
HEIT C, JACKSON B C, MCANDREWS M, et al. Update of the human and mouse serpin gene superfamily[J]. Hum Genomics, 2013, 7(1): 22.
IRVING J A, PIKE R N, DAI W W, et al. Evidence that serpin architecture intrinsically supports papain-like cysteine protease inhibition: engineering α1-antitrypsin to inhibit cathepsin proteases[J]. Biochemistry, 2002, 41(15): 4998-5004.
IKEZOE T. Advances in the diagnosis and treatment of disseminated intravascular coagulation in haematological malignancies[J]. Int J Hematol, 2021, 113(1): 34-44.
FRANÇOIS D, VENISSE L, MARCHAL-SOMME J, et al. Increased expression of protease nexin-1 in fibroblasts during idiopathic pulmonary fibrosis regulates thrombin activity and fibronectin expression[J]. Lab Invest, 2014, 94(11): 1237-1246.
KLIEBER M A, UNDERHILL C, HAMMOND G L, et al. Corticosteroid-binding globulin, a structural basis for steroid transport and proteinase-triggered release[J]. J Biol Chem, 2007, 282(40): 29594-29603.
ZHOU A W, WEI Z Q, READ R J, et al. Structural mechanism for the carriage and release of thyroxine in the blood[J]. Proc Natl Acad Sci U S A, 2006, 103(36): 13321-13326.
ZHANG M, MAGIT D, SAGER R. Expression of maspin in prostate cells is regulated by a positive Ets element and a negative hormonal responsive element site recognized by androgen receptor[J]. Proc Natl Acad Sci U S A, 1997, 94(11): 5673-5678.
HUNT L T, DAYHOFF M O. A surprising new protein superfamily containing ovalbumin, antithrombin-Ⅲ, and α1-proteinase inhibitor[J]. Biochem Biophys Res Commun, 1980, 95(2): 864-871.
TANAKA S, KOGA Y, TAKANO K, et al. Inhibition of chymotrypsin- and subtilisin-like serine proteases with Tk-serpin from hyperthermophilic archaeon Thermococcus kodakaraensis[J]. Biochim Biophys Acta, 2011, 1814(2): 299-307.
KSIAZEK M, MIZGALSKA D, ENGHILD J J, et al. Miropin, a novel bacterial serpin from the periodontopathogen Tannerella forsythia, inhibits a broad range of proteases by using different peptide bonds within the reactive center loop[J]. J Biol Chem, 2015, 290(1): 658-670.
MKAOUAR H, AKERMI N, MARIAULE V, et al. Siropins, novel serine protease inhibitors from gut microbiota acting on human proteases involved in inflammatory bowel diseases[J]. Microb Cell Fact, 2016, 15(1): 201.
NAGY Z A, SZAKÁCS D, BOROS E, et al. Ecotin, a microbial inhibitor of serine proteases, blocks multiple complement dependent and independent microbicidal activities of human serum[J]. PLoS Pathog, 2019, 15(12): e1008232.
WOLLINA U, VOICU C, GIANFALDONI S, et al. Arthrospira platensis-potential in dermatology and beyond[J]. Open Access Maced J Med Sci, 2018, 6(1): 176-180.
AYEHUNIE S, BELAY A, BABA T W, et al. Inhibition of HIV-1 replication by an aqueous extract of Spirulina platensis (Arthrospira platensis)[J]. J Acquir Immune Defic Syndr Hum Retrovirol, 1998, 18(1): 7-12.
ROMAY C H, GONZÁLEZ R, LEDÓN N, et al. C-phycocyanin: a biliprotein with antioxidant, anti-inflammatory and neuroprotective effects[J]. Curr Protein Pept Sci, 2003, 4(3): 207-216.
PORTMANN C, BLOM J F, KAISER M, et al. Isolation of aerucyclamides C and D and structure revision of microcyclamide 7806A: heterocyclic ribosomal peptides from Microcystis aeruginosa PCC 7806 and their antiparasite evaluation[J]. J Nat Prod, 2008, 71(11): 1891-1896.
SYARINA P N, KARTHIVASHAN G, ABAS F, et al. Wound healing potential of Spirulina platensis extracts on human dermal fibroblast cells[J]. EXCLI J, 2015, 14: 385-393.
KWAN J C, EKSIOGLU E A, LIU C, et al. Grassystatins A‒C from marine cyanobacteria, potent cathepsin E inhibitors that reduce antigen presentation[J]. J Med Chem, 2009, 52(18): 5732-5747.
GRAWISH M E. Effects of Spirulina platensis extract on Syrian hamster cheek pouch mucosa painted with 7,12-dimethylbenz[α]anthracene[J]. Oral Oncol, 2008, 44(10): 956-962.
MISHIMA T, MURATA J, TOYOSHIMA M, et al. Inhibition of tumor invasion and metastasis by calcium spirulan (Ca-SP), a novel sulfated polysaccharide derived from a blue-green alga, Spirulina platensis[J]. Clin Exp Metastasis, 1998, 16(6): 541-550.
BRAUNE S, KRÜGER-GENGE A, KAMMERER S, et al. Phycocyanin from Arthrospira platensis as potential anti-cancer drug: review of in vitro and in vivo studies[J]. Life (Basel), 2021, 11(2): 91.
LIU Q, HUANG Y H, ZHANG R H, et al. Medical application of Spirulina platensis derived C-phycocyanin[J]. Evid Based Complement Alternat Med, 2016, 2016: 7803846.
CATASSI A, CESARIO A, ARZANI D, et al. Characterization of apoptosis induced by marine natural products in non small cell lung cancer A549 cells[J]. Cell Mol Life Sci, 2006, 63(19-20): 2377-2386.
MALAKHOV M P, MATTERN M R, MALAKHOVA O A, et al. SUMO fusions and SUMO-specific protease for efficient expression and purification of proteins[J]. J Struct Funct Genomics, 2004, 5(1-2): 75-86.
WEI H D, CAI H Y, WU J W, et al. Heparin binds lamprey angiotensinogen and promotes thrombin inhibition through a template mechanism[J]. J Biol Chem, 2016, 291(48): 24900-24911.
IRVING J A, CABRITA L D, ROSSJOHN J, et al. The 1.5 Å crystal structure of a prokaryote serpin: controlling conformational change in a heated environment[J]. Structure, 2003, 11(4): 387-397.
CABRITA L D, IRVING J A, PEARCE M C, et al. Aeropin from the extremophile Pyrobaculum aerophilum bypasses the serpin misfolding trap[J]. J Biol Chem, 2007, 282(37): 26802-26809.
GOULAS T, KSIAZEK M, GARCIA-FERRER I, et al. A structure-derived snap-trap mechanism of a multispecific serpin from the dysbiotic human oral microbiome[J]. J Biol Chem, 2017, 292(26): 10883-10898.
ANAS A R, NAKAJIMA A, NARUSE C, et al. Determination of FⅦa- sTF inhibitors in toxic Microcystis cyanobacteria by LC-MS technique[J]. Mar Drugs, 2015, 14(1): 7.
WELKER M, BRUNKE M, PREUSSEL K, et al. Diversity and distribution of Microcystis (Cyanobacteria) oligopeptide chemotypes from natural communities studied by single-colony mass spectrometry[J]. Microbiology (Reading), 2004, 150(Pt 6): 1785-1796.
AHMED M N, WAHLSTEN M, JOKELA J, et al. Potent inhibitor of human trypsins from the aeruginosin family of natural products[J]. ACS Chem Biol, 2021, 16(11): 2537-2546.
LIANG J, LI T, ZHANG Y L, et al. Effect of microcystin-LR on protein phosphatase 2A and its function in human amniotic epithelial cells[J]. J Zhejiang Univ Sci B, 2011, 12(12): 951-960.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}