Effect of montelukast on leukotriene B4 metabolism in asthma
HU Yu,1,2,3, XIE Liang1,2,4,5, ZOU Dan3, FU Hongling1,2, LOU Lili1,2, XIE Keqi3, LIU Hanmin,1,2,4,5
1.Department of Pediatric Pulmonology and Immunology, West China Second University Hospital, Sichuan University, Chengdu, 610041, China
2.The Joint Laboratory for Lung Development and Related Diseases of West China Second University Hospital, Sichuan University and School of Life Sciences of Fudan University; West China Institute of Women and Children′s Health, West China Second University Hospital, Sichuan University, Chengdu, 610041, China
3.Department of Pediatrics, Mianyang Central Hospital, School of Medicine, University of Electronic Science and Technology of China, Mianyang, 621000, China
4.Key Laboratory of Birth Defects and Related Diseases of Women and Children (Sichuan University), Ministry of Education, Chengdu, 610041, China
5.NHC Key Laboratory of Chronobiology (Sichuan University), Chengdu, 610041, China
Objective ·To observe the effect of montelukast on the expressions of key genes in LTB4 (leukotriene B4) metabolic pathway in treating asthma and investigate the candidate intervene targets of asthma. Methods ·The acute, subacute, and chronic asthmatic mouse models characterizing by allergic airway disease (AAD) were set up by ovalbumin (OVA) and Al(OH)3 sensitization and challenge and intervened by intragastric administration of montelukast and finally challenged by OVA for chronic asthma model. The pulmonary functions of mice were tested by unconstrained whole body plethysmograph, to quest the change patterns of airway hyperresponsiveness (AHR). The eosinophil (EOS) infiltration and goblet cell (GCL) hyperplasia in mouse lungs were detected by hematoxylin-eosin (HE) staining, to quest the pathologic features of airway allergic inflammation. The levels of immunoglobulin E (IgE), interferon γ (IFN-γ), and interleukin (IL) in bronchoalveolar lavage fluid (BALF) and serum were detected by ELISA and Milliplex kits, to quest the helper T cell type 2 (Th2) inflammation status. The transcription and protein levels of 5-lipoxygenase activating protein (ALOX5AP), leukotriene A4 hydrolase (LTA4H), and leukotriene B4 receptor 1 (BLT1) genes, which encoded the rate-limiting enzymes in LTB4 synthesis pathway, were detected by RT-qPCR, Western blotting and immunohistochemistry (IHC). Results ·The asthmatic mouse model could be set up by OVA and Al (OH)3 and was presented as AHR characterized by increasing enhanced pause (Penh) value, eosinophilic inflammation and high mucous secretion pathologically characterized by airway EOS infiltration and GCL hyperplasia, Th2 inflammation immunologically characterized by the increasing levels of IgE, IL-4, and IL-13 as well as decreasing levels of IFN-γ, IL-2, and IL-12 in BALF and serum. Montelukast could alleviate AAD effectively. The transcription and protein levels of ALOX5AP, LTA4H, and BLT1 genes increased in asthma. Montelukast can inhibit the expression of ALOX5AP gene and promote the expressions of LTB4 and BLT1 genes in asthmatic chronic phase. When challenged by OVA once again, montelukast can induce the significantly high expressions of LTB4 and BLT1 genes. Conclusion ·Montelukast has the effect of relieving allergic inflammation in asthma mice, but it can stimulate the production and accumulation of LTB4 and is significant in chronic phase. When challenged by OVA a second time, LTB4 could be promoted to combine with BLT1 and attend in the pathogenesis of asthma. The results suggested that there was a potential risk of activation of LTB4 by montelukast. The rate-limiting enzyme LTA4H and its receptor BLT1 metabolism may be potential targets for asthma treatment.
HU Yu, XIE Liang, ZOU Dan, FU Hongling, LOU Lili, XIE Keqi, LIU Hanmin. Effect of montelukast on leukotriene B4 metabolism in asthma. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(5): 580-591 doi:10.3969/j.issn.1674-8115.2023.05.008
支气管哮喘是严重威胁人类健康的慢性呼吸系统疾病,全球哮喘患者超过3亿人[1]。哮喘包括过敏性、肥胖性、运动性和药物性等多种表型,其中过敏性哮喘是哮喘的主要表型,多见特异性体质人群在接触变应原后产生的以咳嗽、胸闷、气紧和呼吸困难为主要表现的临床急症。哮喘的病理学特点为气道嗜酸性炎症和黏液高分泌,并以可逆性气流受阻和气道高反应性(airway hyperresponsiveness,AHR)为主要病理生理学特征。过敏性哮喘的发病机制以免疫学改变为特点,主要表现为变应原刺激特异性机体产生Ⅰ型变态反应,导致1型辅助性T细胞(helper T cell type 1,Th1)/Th2比例失衡,Th2增殖活跃并诱导产生大量细胞因子、炎症介质和免疫球蛋白E(immunoglobulin E,IgE),从而促使炎症细胞增殖活化,最终诱发呼吸系统症状。炎症介质在哮喘发病中起关键作用,但其种类繁多且具体作用机制尚不完全清楚。因此,有必要深入探究炎症介质与哮喘发病的潜在联系,拓展其临床治疗思路。
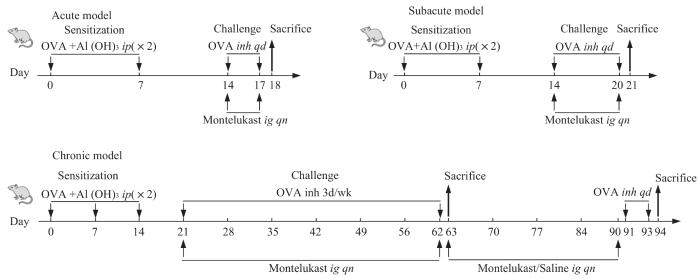
Note: AAD was induced by intraperitoneal sensitization of Balb/c mice with 40 μg grade V OVA and 2 mg aluminum hydroxide in saline. Sensitization was delivered on Day 0 and 14 (acute and subacute models) or on Day 0, 7 and 14 (chronic model). Mice were then challenged by nebulization of 3% OVA in saline for 30 min on four consecutive days (acute), seven consecutive days (subacute), or 3 d/week for 6 weeks (chronic). Control mice were sensitized with 2 mg of aluminum hydroxide in 0.5 mL and challenged by nebulized saline. Treated AAD mice to montelukast by means of gavage every night on challenging days or Day 63 to 90 (chronic). Control mice were treated to gavage saline. AAD mice were challenged by nebulization of 3% OVA in saline for 30 min on three consecutive days by Day 91 to 93.
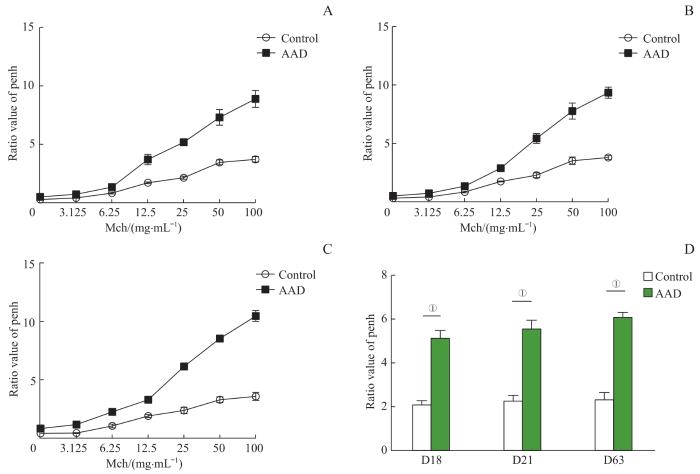
Note: A-C. The Penh values of acute (A), subacute (B) and chronic (C) models of control and AAD mice (n=8). D. The Penh values of each model of control and AAD mice at the concentration of Mch is 25 mg/mL (n=8). ①P=0.000.
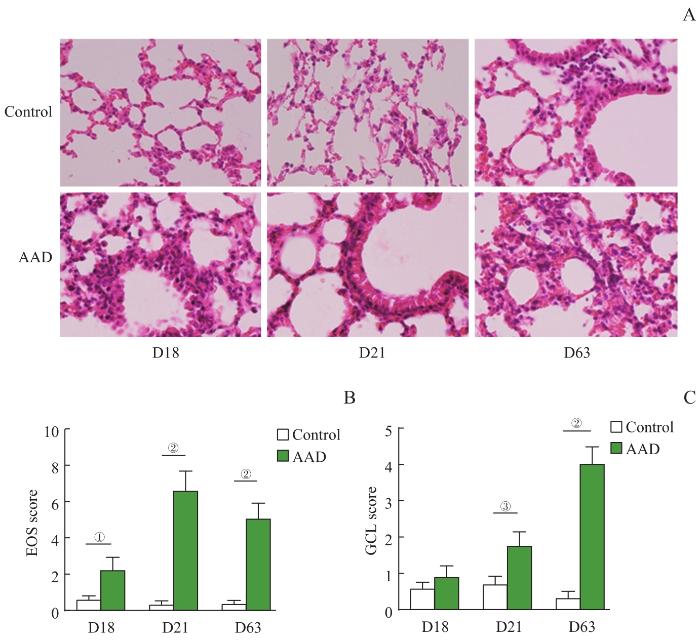
Note: A. The HE staining of acute, subacute and chronic models of control and AAD mice lung samples (×400). B. The EOS scores of control and AAD mice lung samples (n=8). C. The GCL scores of control and AAD mice lung samples (n=8). ①P=0.001, ②P=0.000, ③P=0.003.
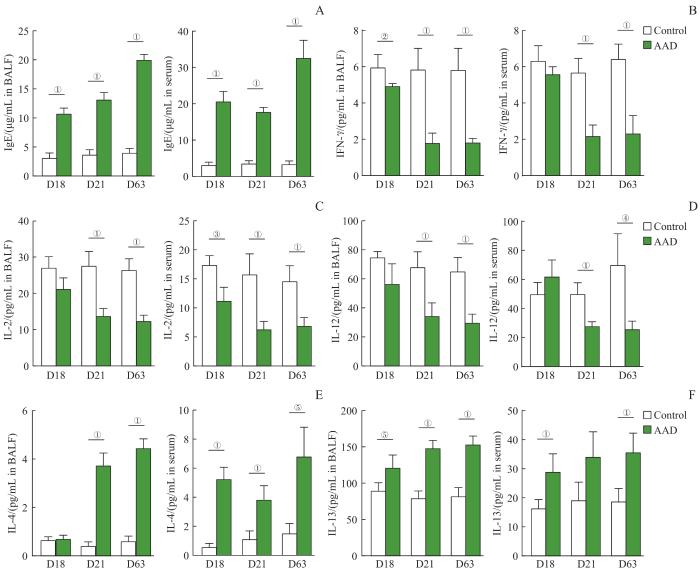
Note: A-F. The expression levels of IgE (A), IFN-γ (B), IL-2 (C), IL-12 (D), IL-4 (E) and IL-13 (F) in BALF and serum of control and AAD mice (n=4). ①P=0.000, ②P=0.004, ③P=0.002, ④P=0.007. ⑤P=0.001, ⑥P=0.006.
Fig 4
Expression levels of IgE and cytokines in BALF and serum of mice
Note: A. Immunohistochemical staining of ALOX5AP protein of control, AAD and AAD+ montelukast mice lung samples (×400). B. RT-qPCR transcription levels of ALOX5AP gene (n=4). C. IOD values of ALOX5AP protein in immunohistochemical staining (n=8). ①P=0.001, ②P=0.000, ③P=0.005.
Fig 5
Transcription and protein levels of ALOX5AP gene
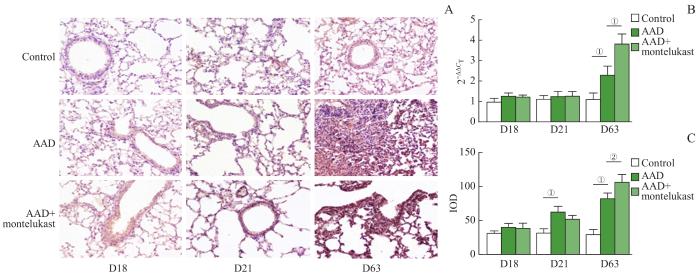
Note: A. Immunohistochemical staining of LTA4H protein of control, AAD and AAD+ montelukast mice lung samples (×400). B. RT-qPCR transcription levels of LTA4H gene (n=4). C. IOD values of LTA4H protein in immunohistochemical staining (n=8). ①P=0.009, ②P=0.000, ③P=0.003.
Fig 6
Transcription and protein levels of LTA4H gene
Note: A. Immunohistochemical staining of BLT1 protein of control, AAD and AAD+montelukast mice lung samples (×400). B. RT-qPCR transcription levels of BLT1 gene (n=4). C.IOD values of BLT1 protein in immunohistochemical staining (n=8). ①P=0.000, ②P=0.003.
Fig 7
Transcription and protein levels of BLT1 gene
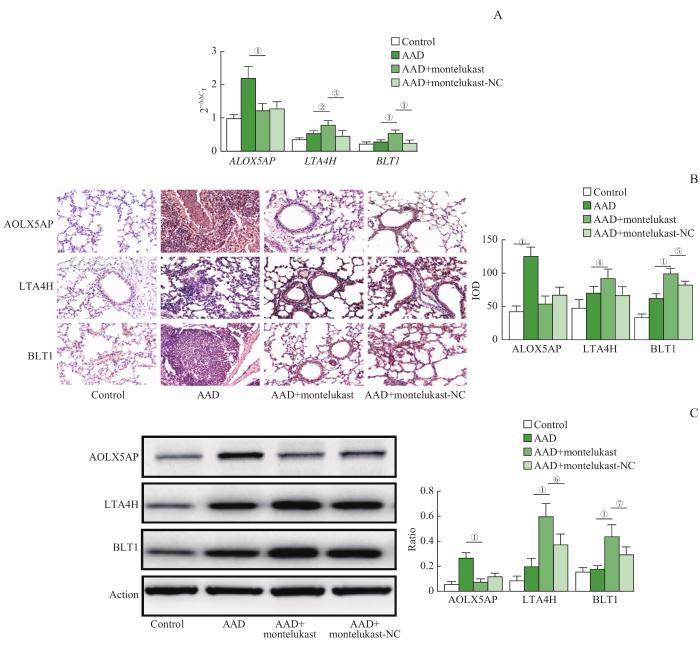
Note: A. RT-qPCR transcription levels of ALOX5AP, LTA4H and BLT1 gene of control, AAD, AAD+montelukast and AAD+montelukast-NC mice (n=4). B. Immunohistochemical staining of ALOX5AP, LTA4H and BLT1 protein lung samples (×400) and the IOD values of each protein in immunohistochemical staining (n=8). C. Western blotting electrophoresis bands of ALOX5AP, LTA4H and BLT1 protein and the ratio values of each protein (n=4). ①P=0.000, ②P=0.007, ③P=0.011, ④P=0.012, ⑤P=0.009, ⑥P=0.017.
Fig 8
Transcription and protein levels of ALOX5AP, LTA4H and BLT1 genes after re-challenged by OVA in asthma mice models
The study was designed by HU Yu and LIU Hanmin. The manuscript was drafted and revised by HU Yu and LIU Hanmin. HU Yu performed the experiments and analyzed the data. XIE Liang, ZOU Dan, FU Hongling, LOU Lili, and XIE Keqi assisted in completing the experiments. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
JOHNSON C C, CHANDRAN A, HAVSTAD S, et al. US childhood asthma incidence rate patterns from the ECHO consortium to identify high-risk groups for primary prevention[J]. JAMA Pediatr, 2021, 175(9): 919-927.
MESHRAM D, BHARDWAJ K, RATHOD C, et al. The role of leukotrienes inhibitors in the management of chronic inflammatory diseases[J]. Recent Pat Inflamm Allergy Drug Discov, 2020, 14(1): 15-31.
YAMAMOTO T, MIYATA J, ARITA M, et al. Current state and future prospect of the therapeutic strategy targeting cysteinyl leukotriene metabolism in asthma[J]. Respir Investig, 2019, 57(6): 534-543.
LOCKE N R, ROYCE S G, WAINEWRIGHT J S, et al. Comparison of airway remodeling in acute, subacute, and chronic models of allergic airways disease[J]. Am J Respir Cell Mol Biol, 2007, 36(5): 625-632.
PADRID P, SNOOK S, FINUCANE T, et al. Persistent airway hyperresponsiveness and histologic alterations after chronic antigen challenge in cats[J]. Am J Respir Crit Care Med, 1995, 151(1): 184-193.
CHO K S, PARK M K, KANG S A, et al. Adipose-derived stem cells ameliorate allergic airway inflammation by inducing regulatory T cells in a mouse model of asthma[J]. Mediators Inflamm, 2014, 2014: 436476.
MIYATA J, FUKUNAGA K, KAWASHIMA Y, et al. Cysteinyl leukotriene metabolism of human eosinophils in allergic disease[J]. Allergol Int, 2020, 69(1): 28-34.
BRUNO F, SPAZIANO G, LIPARULO A, et al. Recent advances in the search for novel 5-lipoxygenase inhibitors for the treatment of asthma[J]. Eur J Med Chem, 2018, 153: 65-72.
LECHNER A, HENKEL F D R, HARTUNG F, et al. Macrophages acquire a TNF-dependent inflammatory memory in allergic asthma[J]. J Allergy Clin Immunol, 2022, 149(6): 2078-2090.
RO M, LEE A J, KIM J H. 5-/ 12-Lipoxygenase-linked cascade contributes to the IL-33-induced synthesis of IL-13 in mast cells, thus promoting asthma development[J]. Allergy, 2018, 73(2): 350-360.
THIVIERGE M, STANKOVÁ J, ROLA-PLESZCZYNSKI M. IL-13 and IL-4 up-regulate cysteinyl leukotriene 1 receptor expression in human monocytes and macrophages[J]. J Immunol, 2001, 167(5): 2855-2860.
ZHOU X J, QIN Z, LU J, et al. Efficacy and safety of salmeterol/fluticasone compared with montelukast alone (or add-on therapy to fluticasone) in the treatment of bronchial asthma in children and adolescents: a systematic review and meta-analysis[J]. Chin Med J, 2021, 134(24): 2954-2961.
WANG W L, LUO X M, ZHANG Q, et al. Bifidobacterium infantis relieves allergic asthma in mice by regulating Th1/Th2[J]. Med Sci Monit, 2020, 26: e920583.
ELIEH ALI KOMI D, BJERMER L. Mast cell-mediated orchestration of the immune responses in human allergic asthma: current insights[J]. Clin Rev Allergy Immunol, 2019, 56(2): 234-247.
BRIGHTLING C E, BRUSSELLE G, ALTMAN P. The impact of the prostaglandin D2 receptor 2 and its downstream effects on the pathophysiology of asthma[J]. Allergy, 2020, 75(4): 761-768.
SCHEXNAYDRE E E, GERSTMEIER J, GARSCHA U, et al. A 5‑lipoxygenase-specific sequence motif impedes enzyme activity and confers dependence on a partner protein[J]. Biochim Biophys Acta Mol Cell Biol Lipids, 2019, 1864(4): 543-551.
MUÑOZ N M, MELITON A Y, MELITON L N, et al. Secretory group V phospholipase A2 regulates acute lung injury and neutrophilic inflammation caused by LPS in mice[J]. Am J Physiol Lung Cell Mol Physiol, 2009, 296(6): L879-L887.
SOKOLOWSKA M, STEFANSKA J, WODZ-NASKIEWICZ K, et al. Cytosolic phospholipase A2 group IVA is overexpressed in patients with persistent asthma and regulated by the promoter microsatellites[J]. J Allergy Clin Immunol, 2010, 125(6): 1393-1395.
GRANATA F, STAIANO R I, LOFFREDO S, et al. The role of mast cell-derived secreted phospholipases A2 in respiratory allergy[J]. Biochimie, 2010, 92(6): 588-593.
TOUQUI L. Antisense inhibition of phospholipase A2: a new approach for already tested therapeutic targets for the treatment of sepsis[J]. Crit Care Med, 2012, 40(7): 2250-2251.
ZAMAN K, HANIGAN M H, SMITH A, et al. Endogenous S-nitrosoglutathione modifies 5-lipoxygenase expression in airway epithelial cells[J]. Am J Respir Cell Mol Biol, 2006, 34(4): 387-393.
PAL K, FENG X, STEINKE J W, et al. Leukotriene A4 hydrolase activation and leukotriene B4 production by eosinophils in severe asthma[J]. Am J Respir Cell Mol Biol, 2019, 60(4): 413-419.
UCHIDA Y, SOMA T, NAKAGOME K, et al. Implications of prostaglandin D2 and leukotrienes in exhaled breath condensates of asthma[J]. Ann Allergy Asthma Immunol, 2019, 123(1): 81-88.e1.
PREEZ S D, RAIDAL S L, DORAN G S, et al. Exhaled breath condensate hydrogen peroxide, pH and leukotriene B4 are associated with lower airway inflammation and airway cytology in the horse[J]. Equine Vet J, 2019, 51(1): 24-32.
STAPLETON R D, SURATT B T, NEFF M J, et al. Bronchoalveolar fluid and plasma inflammatory biomarkers in contemporary ARDS patients[J]. Biomarkers, 2019, 24(4): 352-359.
LEE J J, DIMINA D, MACIAS M P, et al. Defining a link with asthma in mice congenitally deficient in eosinophils[J]. Science, 2004, 305(5691): 1773-1776.
... 支气管哮喘是严重威胁人类健康的慢性呼吸系统疾病,全球哮喘患者超过3亿人[1].哮喘包括过敏性、肥胖性、运动性和药物性等多种表型,其中过敏性哮喘是哮喘的主要表型,多见特异性体质人群在接触变应原后产生的以咳嗽、胸闷、气紧和呼吸困难为主要表现的临床急症.哮喘的病理学特点为气道嗜酸性炎症和黏液高分泌,并以可逆性气流受阻和气道高反应性(airway hyperresponsiveness,AHR)为主要病理生理学特征.过敏性哮喘的发病机制以免疫学改变为特点,主要表现为变应原刺激特异性机体产生Ⅰ型变态反应,导致1型辅助性T细胞(helper T cell type 1,Th1)/Th2比例失衡,Th2增殖活跃并诱导产生大量细胞因子、炎症介质和免疫球蛋白E(immunoglobulin E,IgE),从而促使炎症细胞增殖活化,最终诱发呼吸系统症状.炎症介质在哮喘发病中起关键作用,但其种类繁多且具体作用机制尚不完全清楚.因此,有必要深入探究炎症介质与哮喘发病的潜在联系,拓展其临床治疗思路. ...





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}