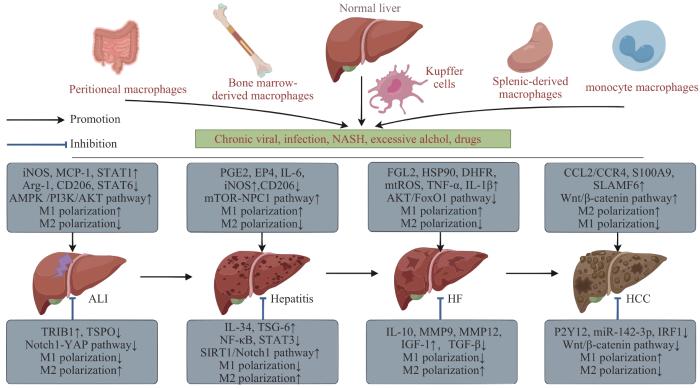
巨噬细胞具有较强的可塑性与异质性,可针对不同信号刺激发生功能转化,如转化为经典激活M1型(即M1型极化)、选择性激活M2型(即M2型极化)等。巨噬细胞M1/M2型极化的途径较为广泛,涉及核因子-κB(nuclear factor-κB,NF-κB)/丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK)信号通路、白细胞介素-4(interleukin-4,IL-4)/信号转导与转录激活因子6(signal transduction and activator of transcription 6,STAT6)信号通路、Notch信号通路、无翼样糖蛋白/β-连环蛋白(Wnt/β-catenin)信号通路等。同时,巨噬细胞M1/M2型极化可不同程度地受到外泌体、代谢物、非编码RNA、电刺激、益生菌等的功能调节,其失衡与不同类型肝病的发生、发展关系密切。该文通过对该极化的作用机制进行梳理,发现巨噬细胞M1型极化在肝组织损伤、炎症反应及纤维化进程中起助推作用,巨噬细胞M2型极化则相反;其中,肝癌作为慢性肝病的晚期阶段,以巨噬细胞M2型极化增强、巨噬细胞M1型极化受损为特征。因此,该文关注巨噬细胞M1/M2型极化在不同类型肝病中的作用,以期能更好地确立巨噬细胞亚群靶向疗法。
关键词:巨噬细胞极化
;
M1型巨噬细胞
;
M2型巨噬细胞
;
肝脏疾病
Abstract
Macrophages have strong plasticity and heterogeneity, and can undergo functional transformation in response to different signal stimuli, such as classical activation of M1 type (M1 type polarization) and selective activation of M2 type (M2 type polarization). The pathways of macrophage M1/M2 polarization are quite extensive, involving nuclear factor-κB (NF-κB)/mitogen-activated protein kinase (MAPK) signaling pathway, interleukin-4 (IL-4)/signal transduction and activator of transcription 6 (STAT6) signaling pathway, Notch signaling pathway, Wnt/β-catenin signaling pathway, etc. At the same time, M1/M2 polarization of macrophages is also regulated by exosomes, metabolites, non-coding RNA, electrical stimulation, probiotics, etc., and its imbalance is closely related to the occurrence and development of different types of liver disease. In this paper, the mechanism of its polarization was reviewed, and it was found that M1 polarization of macrophages played a promoting role in the process of liver tissue injury, inflammation and fibrosis, while M2 polarization of macrophages played the opposite role. Among them, hepatocellular carcinoma, as the advanced stage of chronic liver disease, was characterized by increased M2 polarization and impaired M1 polarization of macrophages. Therefore, this paper pays attention to the role of M1/M2 polarization of macrophages in different types of liver diseases, in order to better establish the targeted therapy of macrophage subsets.
NIU Yuanyuan, WANG Longde, XU Wenjuan, LI Zhengju, ZHANG Ruiting, WU Yuqian. Research progress in the role of M1/M2 polarization of macrophages in different liver diseases. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2024, 44(4): 509-517 doi:10.3969/j.issn.1674-8115.2024.04.012
脂质代谢失调参与非酒精性脂肪性肝炎的病理过程,而胆固醇积累诱导的内质网应激可通过触发巨噬细胞M1型极化来促进该病的发生与发展。如在乙型肝炎病毒X蛋白(hepatitis B virus x protein,HBx)相关的非酒精性脂肪性肝炎中,PEG2通过EP4诱导巨噬细胞M1型极化,进而加速非酒精性脂肪性肝炎的病程进展,其中HBx调节巨噬细胞M1型极化的作用机制与胆固醇由溶酶体向内质网的转运密切相关[37]。
巨噬细胞M1/M2型极化在乙型肝炎病毒(hepatitis B virus,HBV)诱发的损伤修复中起关键作用。LI等[42]研究发现,乙型肝炎表面抗原、乙型肝炎e抗原的分泌增加可通过上调SIRT1的表达、促进Notch1胞内结构域去乙酰化并减少NF-κB核转位,调节由HBV触发的巨噬细胞M1型向M2型的转化,继而提示靶向巨噬细胞中的SIRT1/Notch1途径或可为治疗HBV免疫逃避及慢性感染提供新思路。
NIU Yuanyuan was responsible for collecting and sorting out the literature and writing the main body of the article. WANG Longde conducted the article guidance, review and financial support. XU Wenjuan participated in the literature search. LI Zhengju, ZHANG Ruiting and WU Yuqian participated in the revision of the article and the adjustment of the article frame structure. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
VAN DER HEIDE D, WEISKIRCHEN R, BANSAL R. Therapeutic targeting of hepatic macrophages for the treatment of liver diseases[J]. Front Immunol, 2019, 10: 2852.
XU J, JIN W L, LI X. A new perspective for the treatment of liver fibrosis: targeting macrophage metabolism[J]. Journal of Clinical Hepatology, 2023, 39(4): 922-928.
WANG P, QIU J N, WANG Z X, et al. Research advances in tumor-associated macrophages in hepatocellular carcinoma microenvironment[J]. Journal of Clinical Hepatology, 2023, 39(5): 1212-1218.
SHAPOURI-MOGHADDAM A, MOHAMMADIAN S, VAZINI H, et al. Macrophage plasticity, polarization, and function in health and disease[J]. J Cell Physiol, 2018, 233(9): 6425-6440.
LIU L L, GUO H M, SONG A M, et al. Progranulin inhibits LPS-induced macrophage M1 polarization via NF-κB and MAPK pathways[J]. BMC Immunol, 2020, 21(1): 32.
TAO L R, GUO G, QI Y Y, et al. Inhibition of canonical transient receptor potential 5 channels polarizes macrophages to an M1 phenotype[J]. Pharmacology, 2020, 105(3/4): 202-208.
CHEN W Y, LIU Y N, CHEN J, et al. The Notch signaling pathway regulates macrophage polarization in liver diseases[J]. Int Immunopharmacol, 2021, 99: 107938.
SHANAKI M, KHOSRAVI M, KHOSHDOONI-FARAHANI A, et al. High-intensity interval training reversed high-fat diet-induced M1-macrophage polarization in rat adipose tissue via inhibition of NOTCH signaling[J]. J Inflamm Res, 2020, 13: 165-174.
HU X R, HONG B Z, SHAN X X, et al. The effect of Poria cocos polysaccharide PCP-1C on M1 macrophage polarization via the Notch signaling pathway[J]. Molecules, 2023, 28(5): 2140.
YUAN C, YANG D D, MA J, et al. Modulation of Wnt/β-catenin signaling in IL-17A-mediated macrophage polarization of RAW264.7 cells[J]. Braz J Med Biol Res, 2020, 53(8): e9488.
YANG Y N, ZHAO Z Y, WANG Y P, et al. Effects of hypoxic hepatoma-derived exosome miR-1260b on M2 macrophages and mechanism[J]. Journal of Precision Medicine, 2023, 38(2): 105-110, 115.
WANG Y, GAO R F, LI J P, et al. Downregulation of hsa_circ_0074854 suppresses the migration and invasion in hepatocellular carcinoma via interacting with HuR and via suppressing exosomes-mediated macrophage M2 polarization[J]. Int J Nanomedicine, 2021, 16: 2803-2818.
LI X, LEI Y, WU M, et al. Regulation of macrophage activation and polarization by HCC-derived exosomal lncRNA TUC339[J]. Int J Mol Sci, 2018, 19(10): 2958.
LI D, WANG C Q, MA P F, et al. PGC1α promotes cholangiocarcinoma metastasis by upregulating PDHA1 and MPC1 expression to reverse the Warburg effect[J]. Cell Death Dis, 2018, 9(5): 466.
WANG M, SUN Y, YANG A N, et al. Effect of macrophage PDHA1 gene knockout on apoptosis of hepatocytes in mice with non-alcoholic fatty liver disease[J]. Chinese Journal of Pathophysiology, 2023, 39(1): 123-130.
CHENG J W, CAI W W, ZONG S Y, et al. Metabolite transporters as regulators of macrophage polarization[J]. Naunyn Schmiedeberg's Arch Pharmacol, 2022, 395(1): 13-25.
GHAFOURI-FARD S, ABAK A, TAVAKKOLI AVVAL S, et al. The impact of non-coding RNAs on macrophage polarization[J]. Biomed Pharmacother, 2021, 142: 112112.
LIU D L, WEI Y H, LIU Y D, et al. The long non-coding RNA NEAT1/miR-224-5p/IL-33 axis modulates macrophage M2a polarization and A1 astrocyte activation[J]. Mol Neurobiol, 2021, 58(9): 4506-4519.
WU S Q, XU R, ZHU X, et al. The long noncoding RNA LINC01140/miR-140-5p/FGF9 axis modulates bladder cancer cell aggressiveness and macrophage M2 polarization[J]. Aging, 2020, 12(24): 25845-25864.
WANG Y, LIU H W, ZHAO J S. Macrophage polarization induced by probiotic bacteria: a concise review[J]. Probiotics Antimicrob Proteins, 2020, 12(3): 798-808.
JANG S E, HYAM S R, HAN M J, et al. Lactobacillus brevis G-101 ameliorates colitis in mice by inhibiting NF-κB, MAPK and AKT pathways and by polarizing M1 macrophages to M2-like macrophages[J]. J Appl Microbiol, 2013, 115(3): 888-896.
GU J H, HE X Z, CHEN X Y, et al. Effects of electrical stimulation on cytokine-induced macrophage polarization[J]. J Tissue Eng Regen Med, 2022, 16(5): 448-459.
YANG Y, NI M, ZONG R B, et al. Targeting Notch1-YAP circuit reprograms macrophage polarization and alleviates acute liver injury in mice[J]. Cell Mol Gastroenterol Hepatol, 2023, 15(5): 1085-1104.
WANG Q, WEI S, ZHOU H M, et al. Hyperglycemia exacerbates acetaminophen-induced acute liver injury by promoting liver-resident macrophage proinflammatory response via AMPK/PI3K/AKT-mediated oxidative stress[J]. Cell Death Discov, 2019, 5: 119.
JIN G L, LIU H P, HUANG Y X, et al. Koumine regulates macrophage M1/M2 polarization via TSPO, alleviating sepsis-associated liver injury in mice[J]. Phytomedicine, 2022, 107: 154484.
CHEN Q, SONG Y T, YANG N L, et al. Aging deteriorated liver Ischemia and reperfusion injury by suppressing Tribble's proteins 1 mediated macrophage polarization[J]. Bioengineered, 2022, 13(6): 14519-14533.
LAN Y, QIAN B, HUANG H Y, et al. Hepatocyte-derived prostaglandin E2-modulated macrophage M1-type polarization via mTOR-NPC1 axis-regulated cholesterol transport from lysosomes to the endoplasmic reticulum in hepatitis B virus x protein-related nonalcoholic steatohepatitis[J]. Int J Mol Sci, 2022, 23(19): 11660.
CHI G, PEI J H, LI X Q. EZH2-mediated H3K27me3 promotes autoimmune hepatitis progression by regulating macrophage polarization[J]. Int Immunopharmacol, 2022, 106: 108612.
LIU Y, LIU H, ZHU J S, et al. Interleukin-34 drives macrophage polarization to the M2 phenotype in autoimmune hepatitis[J]. Pathol Res Pract, 2019, 215(8): 152493.
WAN Y M, WU H M, LI Y H, et al. TSG-6 inhibits oxidative stress and induces M2 polarization of hepatic macrophages in mice with alcoholic hepatitis via suppression of STAT3 activation[J]. Front Pharmacol, 2020, 11: 10.
LI J H, YU M X, ZONG R B, et al. Deacetylation of Notch1 by SIRT1 contributes to HBsAg- and HBeAg-mediated M2 macrophage polarization[J]. Am J Physiol Gastrointest Liver Physiol, 2022, 322(4): G459-G471.
LEE C B, KIM M, HAN J, et al. Mesenchymal stem cells influence activation of hepatic stellate cells, and constitute a promising therapy for liver fibrosis[J]. Biomedicines, 2021, 9(11): 1598.
HUANG S S, ZOU Y L, TAN J, et al. CX3CR1 silencing-mediated macrophage polarization attenuates CCl4-induced liver fibrosis in mice[J]. Journal of Army Medical University, 2022, 44(13): 1314-1321.
TAO R, HAN M W, YUAN W, et al. Fibrinogen-like protein 2 promotes proinflammatory macrophage polarization and mitochondrial dysfunction in liver fibrosis[J]. Int Immunopharmacol, 2023, 117: 109631.
PENG Y, LI Z D, CHEN S, et al. DHFR silence alleviated the development of liver fibrosis by affecting the crosstalk between hepatic stellate cells and macrophages[J]. J Cell Mol Med, 2021, 25(21): 10049-10060.
FENG M, DING J, WANG M, et al. Kupffer-derived matrix metalloproteinase-9 contributes to liver fibrosis resolution[J]. Int J Biol Sci, 2018, 14(9): 1033-1040.
RAMACHANDRAN P, PELLICORO A, VERNON M A, et al. Differential Ly-6C expression identifies the recruited macrophage phenotype, which orchestrates the regression of murine liver fibrosis[J]. Proc Natl Acad Sci USA, 2012, 109(46): E3186-E3195.
LV S M, WANG J H, LI L. Extracellular vesicular lncRNA FAL1 promotes hepatocellular carcinoma cell proliferation and invasion by inducing macrophage M2 polarization[J]. J Physiol Biochem, 2023, 79(3): 669-682.
MENG Q, DUAN X Y, YANG Q C, et al. SLAMF6/Ly108 promotes the development of hepatocellular carcinoma via facilitating macrophage M2 polarization[J]. Oncol Lett, 2022, 23(3): 83.
KOHLHEPP M S, LIU H Y, TACKE F, et al. The contradictory roles of macrophages in non-alcoholic fatty liver disease and primary liver cancer: challenges and opportunities[J]. Front Mol Biosci, 2023, 10: 1129831.
YANG L F, ZHANG Z B, WANG L. S100A9 promotes tumor-associated macrophage for M2 macrophage polarization to drive human liver cancer progression: an in vitro study[J]. Kaohsiung J Med Sci, 2023, 39(4): 345-353.
XIE C L, GUO B Q, LIU C Y, et al. Endogenous IFN-β maintains M1 polarization status and inhibits proliferation and invasion of hepatocellular carcinoma cells[J]. Chinese Journal of Cellular and Molecular Immunology, 2016, 32(7): 865-869, 875.
XIE C L, LIU C Y, LIN Y, et al. Effect of IRF1 on polarization and antitumor function of M1 microphage[J]. Basic and Clinical Medicine, 2017, 37(2): 189-196.
PAVLOVIĆ N, KOPSIDA M, GERWINS P, et al. Inhibiting P2Y12 in macrophages induces endoplasmic reticulum stress and promotes an anti-tumoral phenotype[J]. Int J Mol Sci, 2020, 21(21): 8177.
HU Z Q, ZHANG H, LIU W, et al. Mechanism of HBV-positive liver cancer cell exosomal miR-142-3p by inducing ferroptosis of M1 macrophages to promote liver cancer progression[J]. Transl Cancer Res, 2022, 11(5): 1173-1187.
... 脂质代谢失调参与非酒精性脂肪性肝炎的病理过程,而胆固醇积累诱导的内质网应激可通过触发巨噬细胞M1型极化来促进该病的发生与发展.如在乙型肝炎病毒X蛋白(hepatitis B virus x protein,HBx)相关的非酒精性脂肪性肝炎中,PEG2通过EP4诱导巨噬细胞M1型极化,进而加速非酒精性脂肪性肝炎的病程进展,其中HBx调节巨噬细胞M1型极化的作用机制与胆固醇由溶酶体向内质网的转运密切相关[37]. ...
... 巨噬细胞M1/M2型极化在乙型肝炎病毒(hepatitis B virus,HBV)诱发的损伤修复中起关键作用.LI等[42]研究发现,乙型肝炎表面抗原、乙型肝炎e抗原的分泌增加可通过上调SIRT1的表达、促进Notch1胞内结构域去乙酰化并减少NF-κB核转位,调节由HBV触发的巨噬细胞M1型向M2型的转化,继而提示靶向巨噬细胞中的SIRT1/Notch1途径或可为治疗HBV免疫逃避及慢性感染提供新思路. ...



{kind=link}
{kind=link}