Atp2b2 Oblivion heterozygous mutation causes progressive vestibular dysfunction in mice
LIU Yiqing1,2,3, JIN Chenxi1,2,3, FENG Baoyi1,2,3, CHENG Zhenzhe1,2,3, SUN Yilin1,2,3, ZHENG Xiaofei1,2,3, DONG Tingting4, WU Hao1,2,3, TAO Yong,1,2,3
1.Department of Otolaryngology-Head and Neck Surgery, Shanghai Ninth People's Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai 200011, China
2.Ear Institute, Shanghai Jiao Tong University School of Medicine, Shanghai 200125, China
3.Shanghai Key Laboratory of Translation Medicine on Ear and Nose Disease, Shanghai 200125, China
4.Biobank of Shanghai Ninth People's Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai 200125, China
Objective ·To study the alterations in vestibular hair cell morphology and function of ATPase plasma membrane Ca2+ transporting 2 oblivion (Atp2b2 Oblivion) heterozygous mice at different ages. Methods ·Atp2b2 Oblivion heterozygous male mice aged 2 months and 8 months were selected with ten in each kind and C57BL/6J wild-type mice with the same gender, age and number were selected as the control group. Expression patterns of ATP2B2 in vestibular hair cells and numbers of hair cells in the striola zone and the extra striola zone in the two groups of mice at different ages were observed and calculated respectively through immunofluorescence assay. Hair bundle structures were detected by scanning electron microscopy (SEM), and mitochondria and ribbon synapse structures were observed by transmission electron microscopy (TEM). Vestibular evoked potential (VsEP), vestibular evoked myogenic potential (VEMP), rotarod rod test, and balance beam test were adopted for the evaluation of vestibular functions. Results ·ATP2B2 was mainly expressed in the hair bundle of vestibular hair cells in the two groups of mice. Hair cell numbers in the striola zone and the extra-striola zone did not exhibit any differences between Atp2b2 Oblivion heterozygous mutant mice and wild-type mice of 2-month-old and 8-month-old. No visible structural abnormality in the hair bundle could be seen through SEM. TEM results implied no morphological abnormality in mitochondria or ribbon synapses in the 2-month-old heterozygous mutant mice, while vacuolar degeneration was discovered in the mitochondria under the cuticular plate in the 8-month-old heterozygous mutant mice with the normal ribbon synapses and the normal mitochondria near the innervation site. VsEP and VEMP thresholds of 2-month-old and 8-month-old Atp2b2 Oblivion heterozygous mutant mice were significantly elevated compared with the wild-type mice. Analysis of VsEP waveform manifested prolonged P1 latency and declined P1N1 amplitude in heterozygous mutant mice (P<0.05). Results of rotarod rod test and balance beam test acquired from 2-month-old Atp2b2 Oblivion heterozygous mutant mice were not significantly different from the wild-type mice, while the ability of the mutant mice to accomplish the tests descended significantly at 8 months of age compared with the wild-type mice (P<0.05). Conclusion ·Atp2b2 Oblivion heterozygous mutant mice showed defective vestibular electrophysiological function at 2 months old, and abnormalities in vestibule-related behaviors can be detected at 8 months old. The vestibular function of Atp2b2 Oblivion heterozygous mutant mice deteriorate progressively.
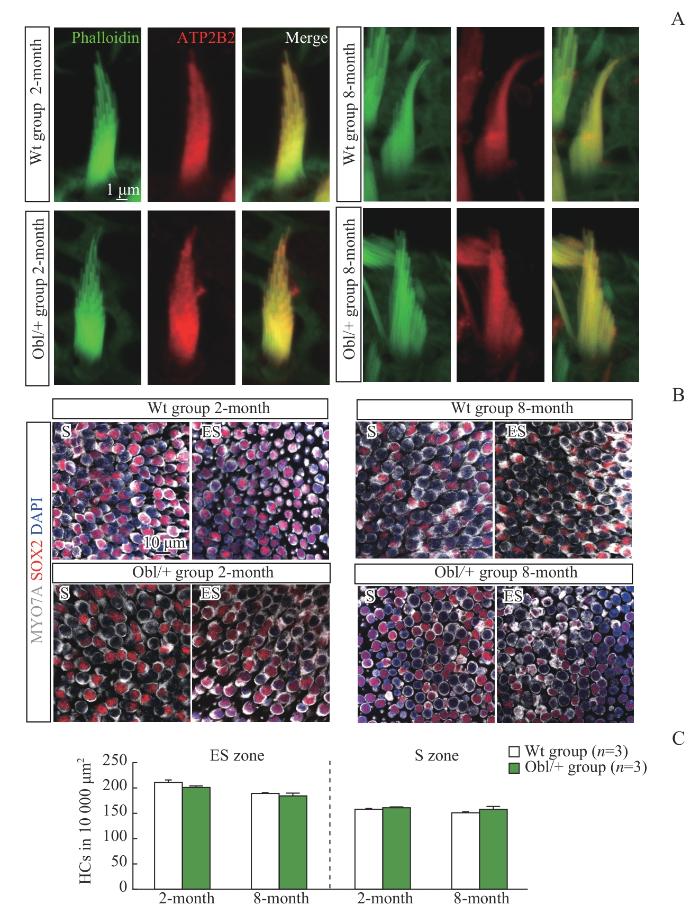
Note: A. Representative confocal images with F-actin (green) and ATP2B2 (red) stained at utricles (×1 000). Scale bar=1 μm. B. Representative confocal images with MYO7A (gray), SOX2 (red) and nuclei (blue) stained at utricles (×126). Scale bar=10 μm. C. Numbers of hair cells (HCs) in an area of 10 000 μm2 in the ES zone and the S zone.
Fig 1
Expression of ATP2B2 and hair cell count in vestibular utricles of the Wt group and the Obl/+ group mice
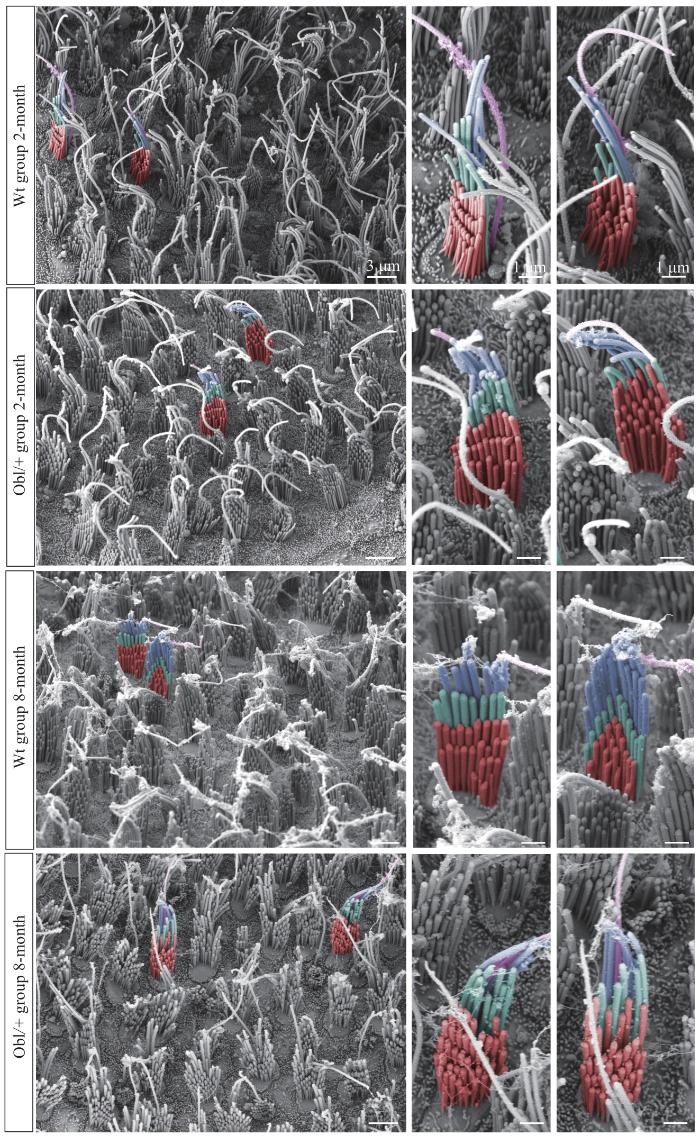
Note: Different colors represent different rows of stereocilia (red—row 3; cyan—row 2; blue—row 1) and violet represents kinocilia. The left column, ×3 000, scale bar=3 μm; the right two columns are partial magnifications of the left column, scale bar=1 μm.
Fig 2
Ultrastructure of hair cell stereocilia in vestibular utricles of the Wt group and the Obl/+ group mice
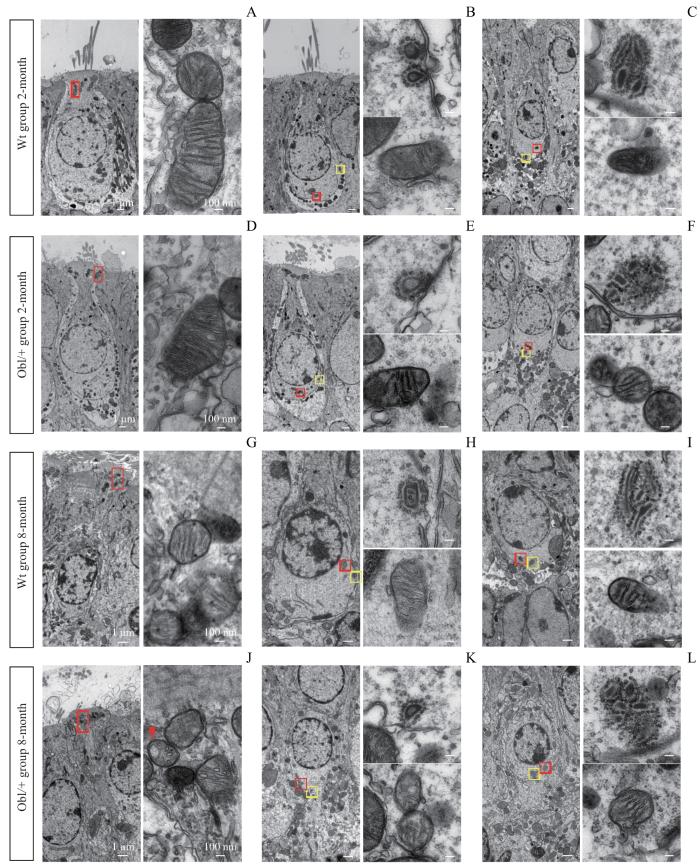
Note: A/D/G/J. Ultrastructure of mitochondria under the cuticular plate in the type Ⅰ utricle hair cells. The images on the right are partial enlargements of the boxes in the left images. Scale bar=1 μm [left: 2-month (×1 500), 8-month (×2 500)] or 100 nm (right: ×15 000). B/C/E/F/H/I/K/L. Ultrastructure of mitochondria and ribbon synapses at the synaptic site in utricle hair cells. The images on the right are partial enlargements of the boxes in the left images. Scale bar=1 μm [left: 2-month (×1 500), 8-month (×2 500)] or 100 nm (right: ×15 000). The red boxes indicate mitochondria; the yellow boxes indicate ribbon synapses; the arrow indicates mitochondrion with vacuole-like denaturation.
Fig 3
Ultrastructure of mitochondria and ribbon synapses of hair cells in utricles of the Wt group and the Obl/+ group mice
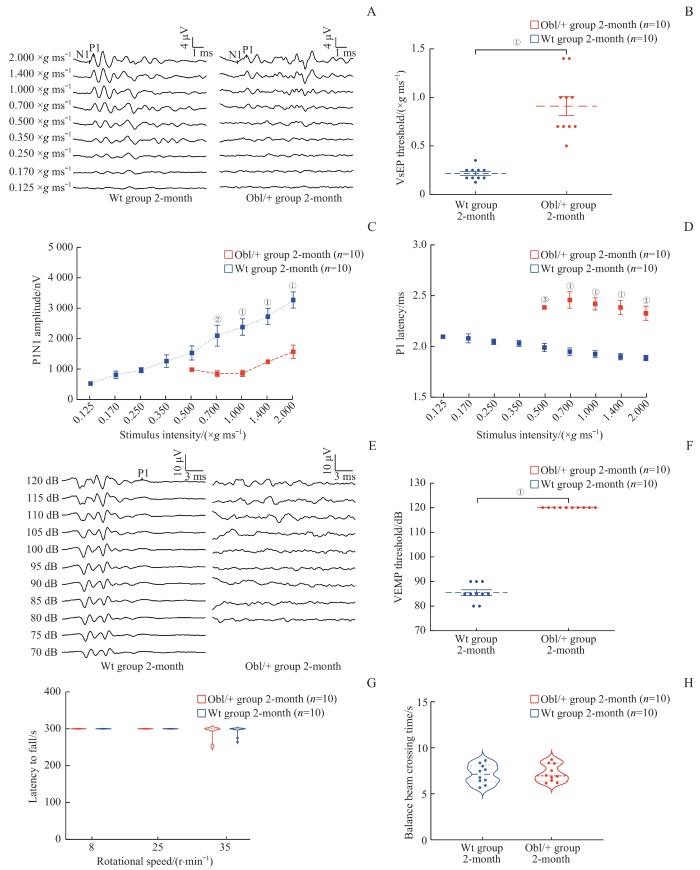
Note: A. VsEP waveform. B. VsEP threshold comparison. C. P1N1 amplitude comparison. D. P1 latency comparison. E. VEMP waveform. F. VEMP threshold comparison. G. Latency to fall from the rotary rod within 300 s. H. Time to pass 80 cm balance beam. ①P=0.000, ②P=0.027, ③P=0.018.
Fig 4
VsEP, VEMP and behavioral tests of 2-month-old mice in the Wt group and the Obl/+ group
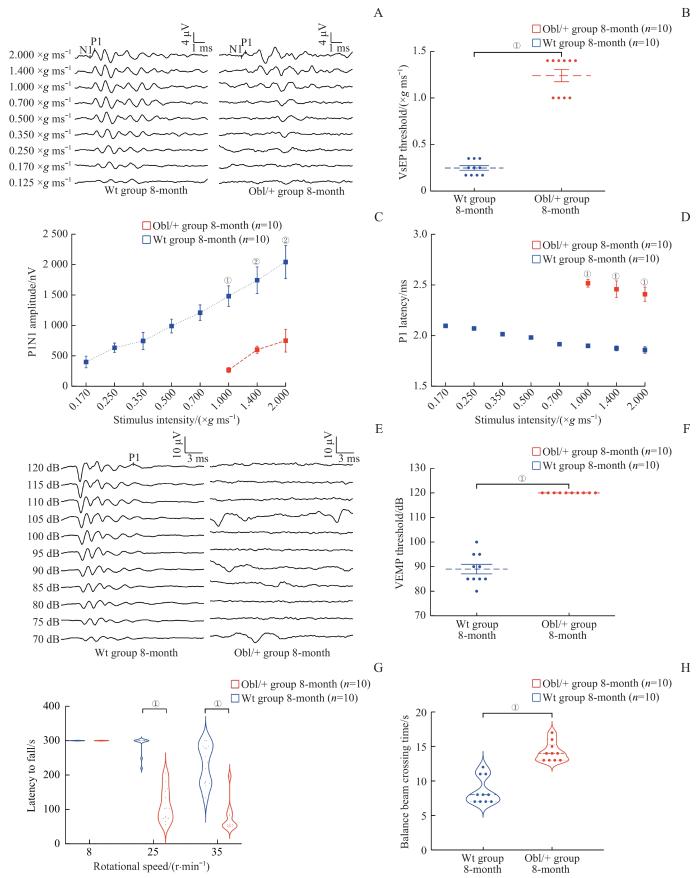
Note:A. VsEP waveform. B. VsEP threshold comparison. C. P1N1 amplitude comparison. D. P1 latency comparison. E. VEMP waveform. F. VEMP threshold comparison. G. Latency to fall from the rotary rod within 300 s. H. Time to pass 80 cm balance beam.①P=0.000, ②P=0.001.
Fig 5
VsEP, VEMP and behavioral tests of 8-month-old mice in the Wt group and the Obl/+ group
LIU Yiqing operated the experiment, acquired and analyzed the data, and drafted the manuscript. JIN Chenxi, FENG Baoyi, CHENG Zhenzhe and SUN Yilin participated in manuscript modification. TAO Yong designed and guided the study and revised the manuscript. ZHENG Xiaofei participated in study designing. Dong Tingting and WU Hao participated in study guidance. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
CHEN Q G, MAHENDRASINGAM S, TICKLE J A, et al. The development, distribution and density of the plasma membrane calcium ATPase 2 calcium pump in rat cochlear hair cells[J]. Eur J Neurosci, 2012, 36(3): 2302-2310.
STAHL W L, EAKIN T J, OWENS J W Jr, et al. Plasma membrane Ca2+-ATPase isoforms: distribution of mRNAs in rat brain by in situ hybridization[J]. Brain Res Mol Brain Res, 1992, 16(3/4): 223-231.
REINHARDT T A, FILOTEO A G, PENNISTON J T, et al. Ca2+-ATPase protein expression in mammary tissue[J]. Am J Physiol Cell Physiol, 2000, 279(5): C1595-C1602.
GREEB J, SHULL G E. Molecular cloning of a third isoform of the calmodulin-sensitive plasma membrane Ca2+-transporting ATPase that is expressed predominantly in brain and skeletal muscle[J]. J Biol Chem, 1989, 264(31): 18569-18576.
KOZEL P J, FRIEDMAN R A, ERWAY L C, et al. Balance and hearing deficits in mice with a null mutation in the gene encoding plasma membrane Ca2+-ATPase isoform 2[J]. J Biol Chem, 1998, 273(30): 18693-18696.
MCCULLOUGH B J, TEMPEL B L. Haplo-insufficiency revealed in deafwaddler mice when tested for hearing loss and ataxia[J]. Hear Res, 2004, 195(1/2): 90-102.
SMITS J J, OOSTRIK J, BEYNON A J, et al. De novo and inherited loss-of-function variants of ATP2B2 are associated with rapidly progressive hearing impairment[J]. Hum Genet, 2019, 138(1): 61-72.
STREET V A, MCKEE-JOHNSON J W, FONSECA R C, et al. Mutations in a plasma membrane Ca2+-ATPase gene cause deafness in deafwaddler mice[J]. Nat Genet, 1998, 19(4): 390-394.
SPECA D J, RABBEE N, CHIHARA D, et al. A genetic screen for behavioral mutations that perturb dopaminergic homeostasis in mice[J]. Genes Brain Behav, 2006, 5(1): 19-28.
WATSON C J, TEMPEL B L. A new Atp2b2 deafwaddler allele, dfwi5, interacts strongly with Cdh23 and other auditory modifiers[J]. Hear Res, 2013, 304: 41-48.
BORTOLOZZI M, BRINI M, PARKINSON N, et al. The novel PMCA2 pump mutation Tommy impairs cytosolic calcium clearance in hair cells and links to deafness in mice[J]. J Biol Chem, 2010, 285(48): 37693-37703.
SPIDEN S L, BORTOLOZZI M, DI LEVA F, et al. The novel mouse mutation Oblivion inactivates the PMCA2 pump and causes progressive hearing loss[J]. PLoS Genet, 2008, 4(10): e1000238.
LEHOTSKÝ J. Plasma membrane Ca2+-pump functional specialization in the brain. Complex of isoform expression and regulation by effectors[J]. Mol Chem Neuropathol, 1995, 25(2/3): 175-187.
TAKAHASHI K, KITAMURA K. A point mutation in a plasma membrane Ca2+-ATPase gene causes deafness in Wriggle Mouse Sagami[J]. Biochem Biophys Res Commun, 1999, 261(3): 773-778.
HRABÉ DE ANGELIS M H, FLASWINKEL H, FUCHS H, et al. Genome-wide, large-scale production of mutant mice by ENU mutagenesis[J]. Nat Genet, 2000, 25(4): 444-447.
HACKNEY C M, MAHENDRASINGAM S, PENN A, et al. The concentrations of calcium buffering proteins in mammalian cochlear hair cells[J]. J Neurosci, 2005, 25(34): 7867-7875.
RICCI A J, WU Y C, FETTIPLACE R. The endogenous calcium buffer and the time course of transducer adaptation in auditory hair cells[J]. J Neurosci, 1998, 18(20): 8261-8277.
FAKIRA A K, GASPERS L D, THOMAS A P, et al. Purkinje cell dysfunction and delayed death in plasma membrane calcium ATPase2-heterozygous mice[J]. Mol Cell Neurosci, 2012, 51(1/2): 22-31.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}