PTEN-regulated alternative splicing of FoxM1 affects tumor cell migration
WANG Xiaoling,1, GE Mengkai2, SHEN Shaoming,2
1.Common Technology Platform, Shanghai Jiao Tong University College of Basic Medical Sciences, Shanghai 200025, China
2.Department of Pathophysiology, Key Laboratory of Cell Differentiation and Apoptosis of Chinese Ministry of Education, Shanghai Jiao Tong University College of Basic Medical Sciences, Shanghai 200025, China
Objective ·To study the effect of phosphatase and tensin homolog on chromosome 10 (PTEN) on alternative splicing of forkhead box M1 (FoxM1), and its impact on tumor cell migration. Methods ·PTEN was knocked down by short hairpin RNA (shRNA) in human embryonic kidney 293T cells, human prostate cancer DU145 cells, human colorectal adenocarcinoma RKO cells, and human colon cancer SW480 and SW620 cells. Specific primers were designed for FoxM1 and its subtypes FoxM1B and FoxM1C, and the mRNA expression levels of FoxM1B and FoxM1C were detected by quantitative real-time PCR (qRT-PCR). FoxM1B and FoxM1C were overexpressed in DU145 cells, and their effects on tumor cell migration were tested by Transwell assay and wound healing assay. Immunofluorescence and dual luciferase reporter gene assay were used to explore the potential mechanism of differential regulation of tumor cell migration by FoxM1B and FoxM1C. Results ·① PTEN was knocked down in 293T, DU145, RKO, SW480, and SW620 cell lines. qRT-PCR results showed that compared with the control cells, the mRNA expression level of FoxM1B significantly increased in PTEN knockdown cells, while the mRNA expression level of FoxM1C decreased or remained unchanged. Knockdown of PTEN did not affect the transcription level of FoxM1, but caused the variable splicing of FoxM1 and promoted the generation of FoxM1B. ② Compared with the control cells, the number of DU145 cells migrating to the below chamber increased in the FoxM1B overexpression group (P=0.024), while the number of migrating DU145 cells in the FoxM1C overexpression group was lower (P=0.000). The healing ability of DU145 cells was significantly enhanced in the FoxM1B overexpression group (P=0.001), while the healing ability of DU145 cells was weakened in the FoxM1C overexpression group (P=0.021). Overall, FoxM1B and FoxM1C had opposite effects on tumor cell migration. FoxM1B promoted tumor cell migration, while FoxM1C inhibited tumor cell migration. ③ Neither FoxM1B nor FoxM1C overexpression could induce β-catenin to enter the nucleus. Dual luciferase reporter gene assay showed no difference in the transcriptional activity of FoxM1B and FoxM1C. The difference between FoxM1B and FoxM1C in the regulation of tumor metastasis was also not mediated by β-catenin translocation. Conclusion ·Knockdown of PTEN regulates the alternative splicing of FoxM1, leading to increasing expression of transcript FoxM1B, which plays a positive role in tumor cell migration.
Keywords:phosphatase and tensin homolog (PTEN)
;
forkhead box M1 (FoxM1)
;
alternative splicing
;
tumor cell
;
migration
WANG Xiaoling, GE Mengkai, SHEN Shaoming. PTEN-regulated alternative splicing of FoxM1 affects tumor cell migration. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(11): 1339-1347 doi:10.3969/j.issn.1674-8115.2023.11.001
10号染色体磷酸酶和张力同源蛋白(phosphatase and tensin homolog,PTEN)是一种重要的抑癌因子[1-2],参与调控细胞的生长、增殖、代谢等多种重要生命活动[3]。PTEN既可作为脂质磷酸酶,也可作为蛋白磷酸酶,主要依赖其双重特异磷酸酶活性起作用。其作为脂质磷酸酶可以通过使底物磷脂酰肌醇3-激酶(phosphatidylinositide 3-kinases,PIP3)去磷酸化,抑制丝氨酸/苏氨酸激酶(serine/threonine kinase,Akt)信号通路,从而发挥抑癌功能[4-5]。此外,PTEN也可以进入细胞核发挥“脚手架”功能,调节细胞周期[6]。近年来,PTEN还被报道与前体mRNA的可变剪接有关[7]。
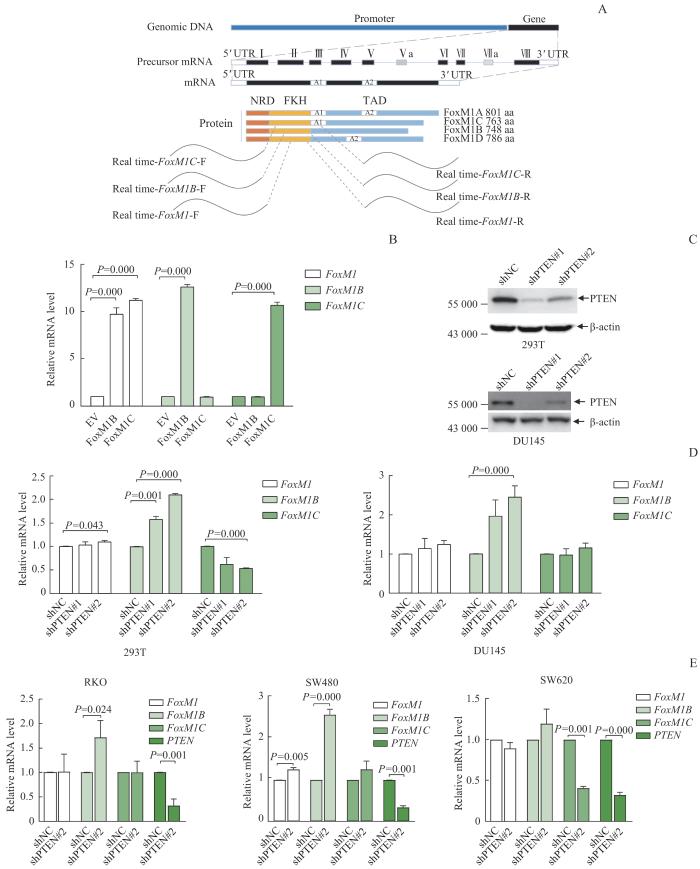
Note: A. Schematic diagram of primer design strategy for mRNA of FoxM1B, FoxM1C and total FoxM1. B. The specificity of the primers was verified by using qRT-PCR in FoxM1B/FoxM1C overexpressing 293T cells. C. Western blotting results showed that PTEN was efficiently knocked down by two pairs of shRNA both in 293T and DU145 cells. D. qRT-PCR results showed that with PTEN knockdown, the expression of FoxM1B mRNA was upregulated in 293T (left) and DU145 (right) cells, whilethe expression of FoxM1C mRNA was downregulated in 293T cells but did not change in DU145 cells. E. qRT-PCR results showed upregulation of FoxM1B mRNA expression, but no significant change in FoxM1C mRNA expression after PTEN being knocked down in RKO (left), SW480 (middle) and SW460 (right) cells. NRD—N-terminal repressor domain; FKH—forkhead/winged-helix domain; TAD—transcativation domain; EV—empty vector.
Fig 1
FoxM1 mRNA isoforms changed in PTEN knockdown cells
Note: A. Western blotting was used to detect the overexpression level of FoxM1B and FoxM1C in DU145 cells. B. Transwell migration analysis results of DU145 cells overexpressing FoxM1B and FoxM1C. Left—typical crystal violet staining photos. Bar=100 μm. Right—statistical analysis of cell migration numbers. C. Wound healing assay of DU145 cells transfected with FoxM1B and FoxM1C overexpression lentivirus. Left—photos of cells incubated for 0 and 24 h in the scratch experiment. Bar=100 μm. Right—statistical analysis of the wound healing rate.
Fig 2
Effects of FoxM1B and FoxM1C on DU145 migration
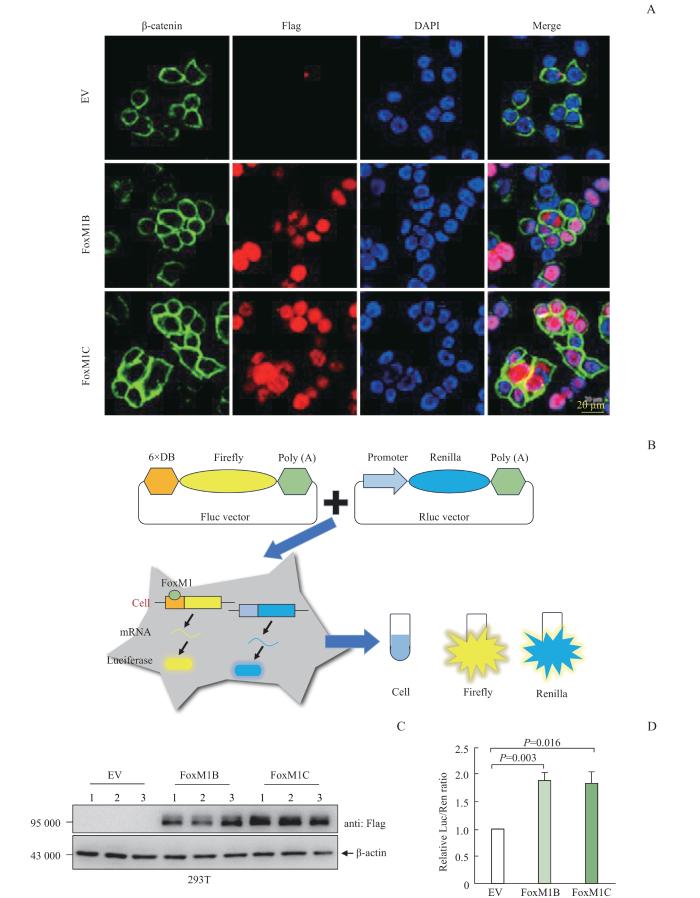
Note: A. Immunofluorescence of 293T cells transfected with Plvx-FOXM1B-Flag and Plvx-FoxM1C-Flag. DAPI represents nucleus, green represents β-catenin, and red represents Flag. Neither FoxM1B nor FoxM1C could cause β-catenin to enter the nucleus. B. Schematic diagram of double luciferase reporter expression system. C. Protein expression of FoxM1B and FoxM1C in FoxM1B/C overexpressing 293T cells transfected with Firefly luciferase and Renilla luciferase reporter gene plasmids. D. Transcriptional activity of FoxM1B and FoxM1C. There was no significant difference in the relative Luc/Ren ratio between FoxM1B and FoxM1C overexpressing 293T cells.
Fig 3
FoxM1B /C transcriptional activities and β-catenin localization
SHEN Shaoming was responsible for project design and thesis revision. GE Mengkai and WANG Xiaoling were responsible for experimental operation, data analysis and thesis writing. All authors have read the final manuscript and agreed to the submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
LIN Y X, WANG Y, DING J X, et al. Reactivation of the tumor suppressor PTEN by mRNA nanoparticles enhances antitumor immunity in preclinical models[J]. Sci Transl Med, 2021, 13(599): eaba9772.
GAO P, HAO J L, XIE Q W, et al. PELO facilitates PLK1-induced the ubiquitination and degradation of Smad4 and promotes the progression of prostate cancer[J]. Oncogene, 2022, 41(21): 2945-2957.
LIU W W, XU L, WANG X, et al. PRDX1 activates autophagy via the PTEN-AKT signaling pathway to protect against cisplatin-induced spiral ganglion neuron damage[J]. Autophagy, 2021, 17(12): 4159-4181.
FENG J W, DANG Y P, ZHANG W Q, et al. PTEN arginine methylation by PRMT6 suppresses PI3K-AKT signaling and modulates pre-mRNA splicing[J]. Proc Natl Acad Sci U S A, 2019, 116(14): 6868-6877.
PENG Q, ZHOU Y J, OYANG L, et al. Impacts and mechanisms of alternative mRNA splicing in cancer metabolism, immune response, and therapeutics[J]. Mol Ther, 2022, 30(3): 1018-1035.
WANG K, DAI X Y, YU A, et al. Peptide-based PROTAC degrader of FOXM1 suppresses cancer and decreases GLUT1 and PD-L1 expression[J]. J Exp Clin Cancer Res, 2022, 41(1): 289.
VANGENDEREN C, HARKNESS T A A, ARNASON T G. The role of anaphase promoting complex activation, inhibition and substrates in cancer development and progression[J]. Aging, 2020, 12(15): 15818-15855.
HU G H, YAN Z W, ZHANG C, et al. FOXM1 promotes hepatocellular carcinoma progression by regulating KIF4A expression[J]. J Exp Clin Cancer Res, 2019, 38(1): 188.
NAKAMURA S, HIRANO I, OKINAKA K, et al. The FOXM1 transcriptional factor promotes the proliferation of leukemia cells through modulation of cell cycle progression in acute myeloid leukemia[J]. Carcinogenesis, 2010, 31(11): 2012-2021.
RATHER T B, PARVEIZ I, BHAT G A, et al. Evaluation of forkhead box M1 (FOXM1) gene expression in colorectal cancer[J]. Clin Exp Med, 2023, 23(6): 2385-2405.
LI S K M, SMITH D K, LEUNG W Y, et al. FoxM1c counteracts oxidative stress-induced senescence and stimulates Bmi-1 expression[J]. J Biol Chem, 2008, 283(24): 16545-16553.
NILSSON M B, SUN H Y, ROBICHAUX J, et al. A YAP/FOXM1 axis mediates EMT-associated EGFR inhibitor resistance and increased expression of spindle assembly checkpoint components[J]. Sci Transl Med, 2020, 12(559): eaaz4589.
SHEN S M, JI Y, ZHANG C, et al. Nuclear PTEN safeguards pre-mRNA splicing to link Golgi apparatus for its tumor suppressive role[J]. Nat Commun, 2018, 9(1): 2392.
MATSUSHITA M, FUJITA K, HAYASHI T, et al. Gut microbiota-derived short-chain fatty acids promote prostate cancer growth via IGF1 signaling[J]. Cancer Res, 2021, 81(15): 4014-4026.
BARGER C J, ZHANG W, HILLMAN J, et al. Genetic determinants of FOXM1 overexpression in epithelial ovarian cancer and functional contribution to cell cycle progression[J]. Oncotarget, 2015, 6(29): 27613-27627.
TASSI R A, TODESCHINI P, SIEGEL E R, et al. FOXM1 expression is significantly associated with chemotherapy resistance and adverse prognosis in non-serous epithelial ovarian cancer patients[J]. J Exp Clin Cancer Res, 2017, 36(1): 63.
BU H T, TANG S S, LIU G T, et al. In silico, in vitro and in vivo studies: dibutyl phthalate promotes prostate cancer cell proliferation by activating Forkhead Box M1 and remission after Natura-α pretreatment[J]. Toxicology, 2023, 488: 153465.
MADHI H, LEE J S, CHOI Y E, et al. FOXM1 inhibition enhances the therapeutic outcome of lung cancer immunotherapy by modulating PD-L1 expression and cell proliferation[J]. Adv Sci (Weinh), 2022, 9(29): e2202702.
HUANG C, XIE D C, CUI J J, et al. FOXM1c promotes pancreatic cancer epithelial-to-mesenchymal transition and metastasis via upregulation of expression of the urokinase plasminogen activator system[J]. Clin Cancer Res, 2014, 20(6): 1477-1488.
LOK G T M, CHAN D W, LIU V W S, et al. Aberrant activation of ERK/FOXM1 signaling cascade triggers the cell migration/invasion in ovarian cancer cells[J]. PLoS One, 2011, 6(8): e23790.
ZHOU Y Z, WANG Q, CHU L, et al. FOXM1c promotes oesophageal cancer metastasis by transcriptionally regulating IRF1 expression[J]. Cell Prolif, 2019, 52(2): e12553.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}