Objective ·To explore the mechanism of changes in resistance to meropenem (MEM), a carbapenem drug, in Acinetobacter baumannii (A. baumannii) cultured in different antibiotic concentrations. Methods ·Through changing the MEM concentration and other culture conditions of the standard sensitive strain of A. baumannii ATCC19606 and the clinical drug-resistant strain AB.2014, the derived strains with different levels of MEM-resistance were induced. The growth curves of all the stains were detected. DNA and RNA of them were extracted. PCR was used to analyze the expression of carbapenemase genes, including IMI, KPC, GES-1, IMP, VIM, NDM-1, OXA23, OXA24, OXA51, and OXA58. Real-time fluorescent quantitative PCR (RT-qPCR) was used to analyze the expression levels of the carbapenemase gene (OXA51), efflux pump genes (adeB, adeG, and adeJ), pore protein genes (carO, omp33-36, and oprC)and the penicillin-binding protein gene (ponA)in the A. baumannii strains with different resistance to MEM, of which the differential gene enrichment was also detected by whole genome sequencing and bioinformatics tools. Results ·The 11 derived strains of ATCC19606 and AB.2014 with different levels of resistance to MEM were obtained, of which the minimum inhibitory concentrations (MIC) were 1‒128 μg/mL. The growth rates and peak values of ATCC19606 and its derivatives decreased with the increase of drug resistance, but AB.2014 and its derivatives did not show this trend. ATCC19606 and its derived strains expressed 3 carbapenemase genes, i.e., OXA51, VIM and IMP, while AB.2014 and most of its derived strains expressed 4 carbapenemase genes, i.e., OXA23, OXA51, VIM and IMP, with only one sensitized derivative of AB.2014 losing OXA23 gene. RT-qPCR results showed that only in ATCC19606 and its drug-resistant derivatives, the expression level of oprC gene decreased with the increase of drug resistance, and the expression levels of most drug-resistant genes were consistent with the changes of drug resistance levels of the strains. Bioinformatics analysis indicated that the differential genes among different ATCC19606 strains were mainly enriched in the siderophore uptake transmembrane transporter activity, cell outer membrane, bacterial secretion system and quorum sensing, while those among AB.2014 strains were mainly enriched in cell outer membrane, cellular response to chemical stimulus, atrazine degradation and RNA polymerase. Conclusion ·The environmental pressure from carbapenems will cause the change of drug resistance of A. baumannii with the involvement of the genes of carbapenemases, efflux pumps, pore proteins and penicillin-binding proteins. The loss of carbapenemase gene OXA23 may lead to the desensitization of drug-resistant A. baumannii strains to carbapenems.
Keywords:Acinetobacter baumannii
;
meropenem (MEM)
;
drug resistance
;
carbapenemase
;
efflux pump
;
pore protein
;
penicillin-binding protein
ZHAO Fumao, PENG Mei, PENG Xiaolu, SHU Weiwei, PENG Li. Changes in drug resistance of Acinetobacter baumannii during the change of meropenem concentration in the environment and its mechanism. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(11): 1396-1407 doi:10.3969/j.issn.1674-8115.2023.11.007
Ex Taq PCR酶预混液(RR001Q)、总RNA提取液(RNAiso Plus,9108)、琼脂糖(5260)购自宝生物工程(大连)有限公司;DNA纯化试剂盒(DP302)、溶菌酶(RT401)购自天根生化科技(北京)有限公司;Evo M-MLV反转录试剂盒(AG11705)、SYBR Green Pro Taq HS预混型试剂盒Ⅱ(AG11741)购自湖南艾科瑞生物工程有限公司;LB培养基(10263/13004)购自青岛日水生物技术有限公司;美罗培南(meropenem,MEM;MB1129)大连美仑生物技术有限公司。DNA文库制备试剂盒(NEBNext® Ultra™ Ⅱ DNA Library Prep Kit for Illumina®,E7645S)购自美国NEB公司。
对ATCC19606、AB.2014及其不同耐药程度的衍生株进行全基因组测序(whole genome sequencing,WGS)。使用DNA文库制备试剂盒(NEB)进行文库构建。在Illumina NextSeq 500平台上,使用配对末端150 bp的序列对汇集的文库进行测序。原始数据的质量由FastQC(v0.11.8)进行评估,然后使用Trimomatic(v0.39)[18]进行质量剪切,以获得相对准确和有效的数据。参考GATK(v4.1.1.0)[19]最佳实践推荐的设计过程,使用BWA(v0.7.17)将样本的有效数据与参考基因组进行比较(使用ATCC19606作为参考基因组,登录号为NZ_CP043953.1)。比较结果通过SAMtools(v1.9)进行格式转换和排序,使用GATK的重复标记来标记重复序列。根据比较结果进行冗余序列分析和插入片段分布分析。BEDTools(v2.28.0)用于深度覆盖率的统计分析。使用GATK的单倍型调用者分析每个样本和参考基因组之间的基因型差异,合并和整合不同样本的分析,以获得样本的变异信息。SnpEff(v4.3T)用于根据参考基因组的注释信息注释突变。使用R语言包topGO进行GO(Gene Ontology)富集分析。使用R语言包clusterProfiler[20]进行KEGG(Kyoto Encyclopedia of Genes and Genomes)路径和COG(Clusters of Orthologous Groups of Proteins)分类富集分析。
Note: A. Detection of OXA23 gene expression in ATCC19606 and its MEM-resistant variants. B.Detection of OXA23 gene expression in AB.2014 and its MEM-resistant variants. C.Detection of OXA23 gene expression in MEM-sensitive variants.
Fig 2
Analysis of OXA23 gene expression in A. baumannii strains with different drug resistance by PCR
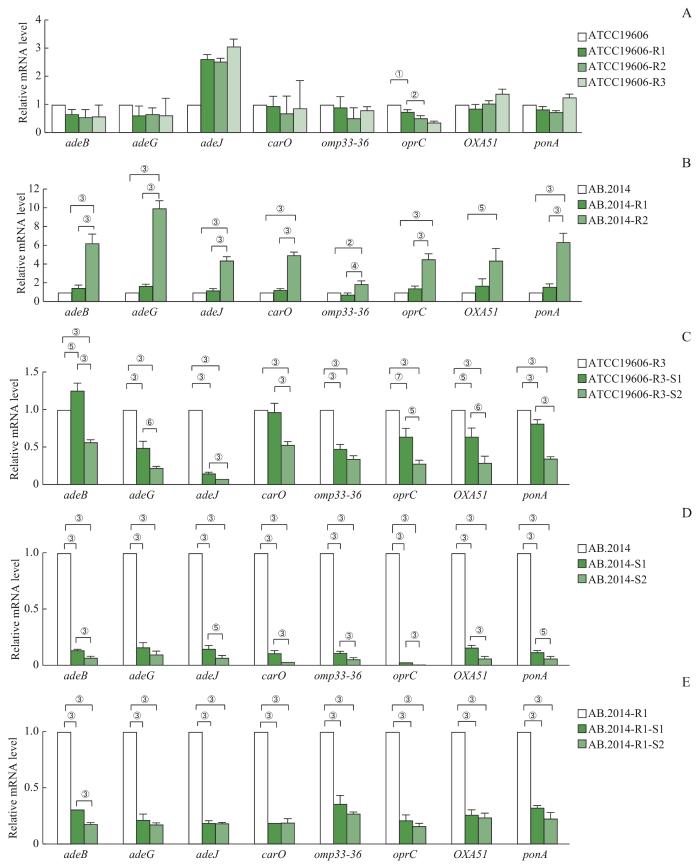
Note: A. Expression profiles of the genes in ATCC19606 and its MEM-resistant variants. B. Expression profiles of the genes in AB.2014 and its MEM-resistant variants. C. Expression profiles of the genes in ATCC19606-R3 and its MEM-sensitive variants. D. Expression profiles of the genes in AB.2014 and its MEM-sensitive variants. E. Expression profiles of the genes of AB.2014-R1 and its MEM-sensitive variants. ①P=0.005, ②P= 0.003, ③P=0.000, ④P=0.007, ⑤P=0.001, ⑥P=0.002, ⑦P=0.006.
Fig 3
RT-qPCR analysis of MEM-resistance-related genes in A. baumannii strains with different drug resistance
Note: A. Enrichment of differential genes in ATCC19606 compared with ATCC19606-R3. B. Enrichment of differential genes in ATCC19606-R3-S2 compared with ATCC19606-R3. C. Enrichment of differential genes in AB.2014 compared with AB.2014-R3. D. Enrichment of differential genes in AB.2014-R3 compared with AB.2014. E. Enrichment of differential genes in AB.2014 compared with AB.2014-S2. F. Enrichment of differential genes in AB.2014-S2 compared with AB.2014. G. Enrichment of differential genes in ATCC19606 compared with AB.2014. H. Enrichment of differential genes in AB.2014 compared with ATCC19606.
Fig 4
GO analysis of differential genes among ATCC19606, AB.2014 and their derivatives
Note: A. Enrichment of differential genes in ATCC19606 compared with ATCC19606-R3. B. Enrichment of differential genes in ATCC19606-R3 compared with ATCC19606. C. Enrichment of differential genes in ATCC19606-R3 compared with ATCC19606-R3-S2. D. Enrichment of differential genes in AB.2014 compared with AB.2014-R3. E. Enrichment of differential genes in AB.2014 compared with AB.2014-S2. F. Enrichment of differential genes in AB.2014-R2-S2 compared with AB.2014-R2.
Fig 5
KEGG analysis of differential genes among ATCC19606, AB.2014 and their derivatives
The study was designed by ZHAO Fumao and PENG Li. The experimental operations were completed by ZHAO Fumao, PENG Mei, PENG Xiaolu and SHU Weiwei. The results were analyzed by ZHAO Fumao. The manuscript was drafted and revised by ZHAO Fumao, PENG Mei and PENG Li. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
DOURAGHI M, KENYON J J, ARIS P, et al. Accumulation of antibiotic resistance genes in carbapenem-resistant Acinetobacter baumannii isolates belonging to lineage 2, global clone 1, from outbreaks in 2012-2013 at a Tehran burns hospital[J]. mSphere, 2020, 5(2): e00164-e00120.
RAMIREZ M S, BONOMO R A, TOLMASKY M E. Carbapenemases: transforming Acinetobacter baumannii into a yet more dangerous menace[J]. Biomolecules, 2020, 10(5): 720.
MENG X, FU J T, ZHENG Y, et al. Ten-year changes in bloodstream infection with Acinetobacter baumannii complex in intensive care units in Eastern China: a retrospective cohort study[J]. Front Med (Lausanne), 2021, 8: 715213.
HAMIDIAN M, NIGRO S J. Emergence, molecular mechanisms and global spread of carbapenem-resistant Acinetobacter baumannii[J]. Microb Genom, 2019, 5(10): e000306.
CHOQUET M, LOHOU E, PAIR E, et al. Efflux pump overexpression profiling in Acinetobacter baumannii and study of new 1-(1-naphthylmethyl)-piperazine analogs as potential efflux inhibitors[J]. Antimicrob Agents Chemother, 2021, 65(9): e0071021.
UPPALAPATI S R, SETT A, PATHANIA R. The outer membrane proteins OmpA, CarO, and OprD of Acinetobacter baumannii confer a two-pronged defense in facilitating its success as a potent human pathogen[J]. Front Microbiol, 2020, 11: 589234.
HERNÁNDEZ-ROCAMORA V M, BARANOVA N, PETERS K, et al. Real-time monitoring of peptidoglycan synthesis by membrane-reconstituted penicillin-binding proteins[J]. eLife, 2021, 10: e61525.
BOLL J M, CROFTS A A, PETERS K, et al. A penicillin-binding protein inhibits selection of colistin-resistant, lipooligosaccharide-deficient Acinetobacter baumannii[J]. Proc Natl Acad Sci USA, 2016, 113(41): E6228-E6237.
MOSTAFAVI S N, KHEDMATI M, KELISHADI R. Microbiology and antimicrobial sensitivity of ventriculo-peritoneal shunt infections in a referral paediatric neurosurgery ward during a period of 7 years[J]. J Glob Antimicrob Resist, 2022, 29: 63-67.
CHEN X, MENG X B, GAO Q Q, et al. Meropenem selection induced overproduction of the intrinsic carbapenemase as well as phenotype divergence in Acinetobacter baumannii[J]. Int J Antimicrob Agents, 2017, 50(3): 419-426.
SHAHCHERAGHI F, NIKBIN V S, FEIZABADI M M. Prevalence of ESBLs genes among multidrug-resistant isolates of Pseudomonas aeruginosa isolated from patients in Tehran[J]. Microb Drug Resist, 2009, 15(1): 37-39.
KUO H Y, CHANG K C, KUO J W, et al. Imipenem: a potent inducer of multidrug resistance in Acinetobacter baumannii[J]. Int J Antimicrob Agents, 2012, 39(1): 33-38.
HU W S, YAO S M, FUNG C P, et al. An OXA-66/OXA-51-like carbapenemase and possibly an efflux pump are associated with resistance to imipenem in Acinetobacter baumannii[J]. Antimicrob Agents Chemother, 2007, 51(11): 3844-3852.
RUMBO C, GATO E, LÓPEZ M, et al. Contribution of efflux pumps, porins, and β-lactamases to multidrug resistance in clinical isolates of Acinetobacter baumannii[J]. Antimicrob Agents Chemother, 2013, 57(11): 5247-5257.
SEWE S O, SILVA G, SICAT P, et al. Trimming and validation of illumina short reads using trimmomatic, trinity assembly, and assessment of RNA-seq data[J]. Methods Mol Biol, 2022, 2443: 211-232.
KNOPP M, ANDERSSON D I. Amelioration of the fitness costs of antibiotic resistance due to reduced outer membrane permeability by upregulation of alternative porins[J]. Mol Biol Evol, 2015, 32(12): 3252-3263.
DA SILVA K E, MACIEL W G, CRODA J, et al. A high mortality rate associated with multidrug-resistant Acinetobacter baumannii ST79 and ST25 carrying OXA-23 in a Brazilian intensive care unit[J]. PLoS One, 2018, 13(12): e0209367.
BLACKWELL G A, HALL R M. Mobilisation of a small Acinetobacter plasmid carrying an oriT transfer origin by conjugative RepAci6 plasmids[J]. Plasmid, 2019, 103: 36-44.
BARTSCH A, IVES C M, KATTNER C, et al. An antibiotic-resistance conferring mutation in a neisserial porin: structure, ion flux, and ampicillin binding[J]. Biochim Biophys Acta Biomembr, 2021, 1863(6): 183601.
BHAMIDIMARRI S P, YOUNG T R, SHANMUGAM M, et al. Acquisition of ionic copper by the bacterial outer membrane protein OprC through a novel binding site[J]. PLoS Biol, 2021, 19(11): e3001446.
ZHANG M L, CHEN L H, YE C S, et al. Co-selection of antibiotic resistance via copper shock loading on bacteria from a drinking water bio-filter[J]. Environ Pollut, 2018, 233: 132-141.
SCOFFONE V C, TRESPIDI G, BARBIERI G, et al. Role of RND efflux pumps in drug resistance of cystic fibrosis pathogens[J]. Antibiotics (Basel), 2021, 10(7): 863.
TOTH M, LEE M, STEWART N K, et al. Effects of inactivation of D,D-transpeptidases of Acinetobacter baumannii on bacterial growth and susceptibility to β-lactam antibiotics[J]. Antimicrob Agents Chemother, 2022, 66(1): e0172921.
YI L, DONG X, GRENIER D, et al. Research progress of bacterial quorum sensing receptors: classification, structure, function and characteristics[J]. Sci Total Environ, 2021, 763: 143031.
VALASTYAN J S, KRAML C M, PELCZER I, et al. Saccharomyces cerevisiae requires CFF1 to produce 4-hydroxy-5-methylfuran-3(2H)- one, a mimic of the bacterial quorum-sensing autoinducer AI-2[J]. mBio, 2021, 12(2): e03303-e03320.
... 对ATCC19606、AB.2014及其不同耐药程度的衍生株进行全基因组测序(whole genome sequencing,WGS).使用DNA文库制备试剂盒(NEB)进行文库构建.在Illumina NextSeq 500平台上,使用配对末端150 bp的序列对汇集的文库进行测序.原始数据的质量由FastQC(v0.11.8)进行评估,然后使用Trimomatic(v0.39)[18]进行质量剪切,以获得相对准确和有效的数据.参考GATK(v4.1.1.0)[19]最佳实践推荐的设计过程,使用BWA(v0.7.17)将样本的有效数据与参考基因组进行比较(使用ATCC19606作为参考基因组,登录号为NZ_CP043953.1).比较结果通过SAMtools(v1.9)进行格式转换和排序,使用GATK的重复标记来标记重复序列.根据比较结果进行冗余序列分析和插入片段分布分析.BEDTools(v2.28.0)用于深度覆盖率的统计分析.使用GATK的单倍型调用者分析每个样本和参考基因组之间的基因型差异,合并和整合不同样本的分析,以获得样本的变异信息.SnpEff(v4.3T)用于根据参考基因组的注释信息注释突变.使用R语言包topGO进行GO(Gene Ontology)富集分析.使用R语言包clusterProfiler[20]进行KEGG(Kyoto Encyclopedia of Genes and Genomes)路径和COG(Clusters of Orthologous Groups of Proteins)分类富集分析. ...
1
... 对ATCC19606、AB.2014及其不同耐药程度的衍生株进行全基因组测序(whole genome sequencing,WGS).使用DNA文库制备试剂盒(NEB)进行文库构建.在Illumina NextSeq 500平台上,使用配对末端150 bp的序列对汇集的文库进行测序.原始数据的质量由FastQC(v0.11.8)进行评估,然后使用Trimomatic(v0.39)[18]进行质量剪切,以获得相对准确和有效的数据.参考GATK(v4.1.1.0)[19]最佳实践推荐的设计过程,使用BWA(v0.7.17)将样本的有效数据与参考基因组进行比较(使用ATCC19606作为参考基因组,登录号为NZ_CP043953.1).比较结果通过SAMtools(v1.9)进行格式转换和排序,使用GATK的重复标记来标记重复序列.根据比较结果进行冗余序列分析和插入片段分布分析.BEDTools(v2.28.0)用于深度覆盖率的统计分析.使用GATK的单倍型调用者分析每个样本和参考基因组之间的基因型差异,合并和整合不同样本的分析,以获得样本的变异信息.SnpEff(v4.3T)用于根据参考基因组的注释信息注释突变.使用R语言包topGO进行GO(Gene Ontology)富集分析.使用R语言包clusterProfiler[20]进行KEGG(Kyoto Encyclopedia of Genes and Genomes)路径和COG(Clusters of Orthologous Groups of Proteins)分类富集分析. ...
1
... 对ATCC19606、AB.2014及其不同耐药程度的衍生株进行全基因组测序(whole genome sequencing,WGS).使用DNA文库制备试剂盒(NEB)进行文库构建.在Illumina NextSeq 500平台上,使用配对末端150 bp的序列对汇集的文库进行测序.原始数据的质量由FastQC(v0.11.8)进行评估,然后使用Trimomatic(v0.39)[18]进行质量剪切,以获得相对准确和有效的数据.参考GATK(v4.1.1.0)[19]最佳实践推荐的设计过程,使用BWA(v0.7.17)将样本的有效数据与参考基因组进行比较(使用ATCC19606作为参考基因组,登录号为NZ_CP043953.1).比较结果通过SAMtools(v1.9)进行格式转换和排序,使用GATK的重复标记来标记重复序列.根据比较结果进行冗余序列分析和插入片段分布分析.BEDTools(v2.28.0)用于深度覆盖率的统计分析.使用GATK的单倍型调用者分析每个样本和参考基因组之间的基因型差异,合并和整合不同样本的分析,以获得样本的变异信息.SnpEff(v4.3T)用于根据参考基因组的注释信息注释突变.使用R语言包topGO进行GO(Gene Ontology)富集分析.使用R语言包clusterProfiler[20]进行KEGG(Kyoto Encyclopedia of Genes and Genomes)路径和COG(Clusters of Orthologous Groups of Proteins)分类富集分析. ...







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}