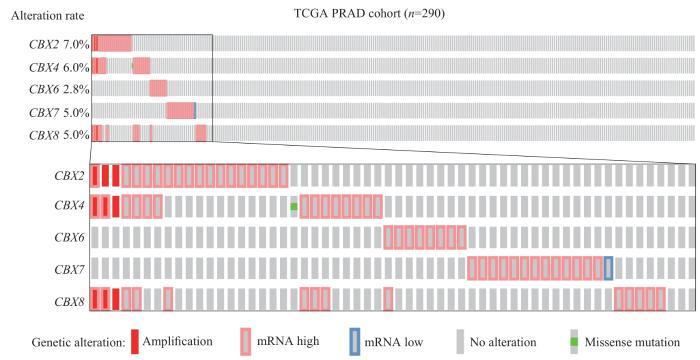
目的·探究染色质组织调节框同源蛋白8(chromobox protein homolog 8,CBX8)在前列腺癌中的生物学功能,并通过转录组及表观修饰分析揭示CBX8在前列腺癌转移中的作用机制。方法·利用cBioPortal数据库对癌症基因组图谱(The Cancer Genome Atlas,TCGA)中前列腺腺癌(prostate adenocarcinoma,PRAD)患者样本数据集进行CBX家族蛋白mRNA表达分析。采用短发夹RNA技术敲低DU145前列腺癌细胞系中的CBX8,通过CCK-8和Transwell实验检测细胞增殖和侵袭水平的变化。使用RNA转录组测序(RNA-seq)分析敲低CBX8后影响的差异表达基因。对这些差异表达基因进行基因集富集分析(Gene Set Enrichment Analysis,GSEA)、基因本体论(Gene Ontology,GO)功能分析以及京都基因和基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)信号通路富集分析。通过染色质免疫沉淀测序(ChIP-seq)观察和测定敲低CBX8后基因组H3K27me3甲基化水平的变化。结果·根据对TCGA-PRAD患者样本数据的分析,发现CBX8 mRNA在前列腺癌中高表达。在前列腺癌细胞系DU145中敲低CBX8后,细胞的增殖能力没有显著变化(P>0.05),但其侵袭能力却显著提高(P<0.05)。RNA-seq分析显示CBX8敲低导致750个基因表达上调,951个基因表达下调;其中,与多种肿瘤转移有关的支链氨基酸转氨酶1(branched-chain-amino-acid aminotransferase 1,BCAT1)在敲除CBX8后表达明显上升。GSEA显示表达水平受影响的基因与多梳蛋白复合体1(polycomb repressive complex 1,PRC1)的功能有关。同时,通过GO和KEGG信号通路富集分析发现受影响的生物过程包括转运RNA(transfer RNA,tRNA)氨酰化、DNA复制、氨酰基-tRNA连接酶活性变化以及钙黏蛋白的结合等;特别是在GO功能分析的细胞组分方面富集了与肿瘤转移有关的细胞-基底连接相关基因。利用ChIP-seq对表观修饰的研究显示,在敲低CBX8后全基因组的H3K27me3水平有所下降;并鉴定了97个位于CBX8敲低后转录上调基因附近的位点,其中包括BCAT1转录起始位点。结论·CBX8在人前列腺癌中高表达。CBX8具有抑制肿瘤细胞侵袭的功能。其机制可能是CBX8/PRC1复合体结合于BCAT1转录起始位点并抑制BCAT1转录。
关键词:染色质组织调节框同源蛋白8
;
前列腺癌
;
细胞增殖
;
肿瘤侵袭
Abstract
Objective ·To elucidate the regulatory mechanisms of the chromobox protein homolog 8 (CBX8) in prostate cancer metastasis from transcriptome and epigenetic modification perspectives. Methods ·The correlation between the expression of CBX proteins and prostate adenocarcinoma (PRAD) in The Cancer Genome Atlas (TCGA) was examined through an analysis based on cBioPortal database. A stable CBX8 knockdown DU145 prostate cancer cell line was established via short hairpin RNA (shRNA) transfection. Subsequently, the proliferation and invasion of the CBX8 knockdown cells were analyzed by CCK-8 assay and Transwell assay, respectively. Transcriptome changes of the CBX8 knockdown cells were investigated through RNA sequencing (RNA-seq) coupled with Gene Set Enrichment Analysis (GSEA). To further evaluate the functional implications of these transcriptomic alterations, Gene Ontology (GO) for functional analysis was deployed. Moreover, to identify potentially affected signalling pathways, the Kyoto Encyclopedia of Genes and Genomes (KEGG) was utilized for pathway enrichment analysis. Lastly, the levels of H3K27me3, a key histone modification associated with CBX8, in the knockdown cells were determined by chromatin immunoprecipitation sequencing (ChIP-seq). Results ·Bioinformatic analysis with cBioPortal database, based on TCGA-PRAD cohorts, unveiled a high CBX8 mRNA expression in PRAD. Knockdown of CBX8 did not significantly affect the proliferation of DU145 cells (P>0.05), but caused a a significant increase in their invasiveness (P<0.05). The RNA-seq analysis revealed that CBX8 knockdown led to the upregulation of 750 genes and the downregulation of 951 genes. Notably, branched-chain-amino-acid aminotransferase 1 (BCAT1), a gene implicated in the metastasis of various types of cancers, showed a significant increase in expression following CBX8 knockdown. GSEA showed that the expression levels were of the affected genes were related to the functions of the polycomb repressive complex 1 (PRC1). A further investigation using GO and KEGG analyses identified several enriched pathways in the CBX8 knockdown cells, including transfer RNA (tRNA) aminoacylation, DNA replication, changes in aminoacyl-tRNA ligase activity, and cadherin binding. Interestingly, in terms of cell component of GO functional analysis, cell-substrate junction-related genes associated with tumor metastasis appeared to be enriched. ChIP-seq results showed a global decrease in H3K27me3 levels. Significantly, 97 reduced H3K27me3 peaks were found located nearby genes that were upregulated upon CBX8 knockdown, including the transcriptional start site of BCAT1. Conclusion ·CBX8 is highly expressed in prostate cancer. CBX8 suppresses prostate cancer cell invasion, possibly by recruiting the transcriptional repressive PRC1 complex to the transcription site of BCAT1, thereby inhibiting BCAT1 transcription and tumor metastasis.
Keywords:chromobox protein homolog 8 (CBX8)
;
prostate cancer
;
cell proliferation
;
tumor invasion
YANG Wanli, SONG Juan, LI Bing, LAO Yimin. Deciphering the suppressive effects of CBX8 on prostate cancer cell invasion. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(12): 1507-1519 doi:10.3969/j.issn.1674-8115.2023.12.005
CBX8稳定敲低细胞株与对照细胞株抽提RNA后由北京安诺优达基因科技公司进行RNA转录组测序(RNA-seq)建库和测序。用带有Oligo(dT)的磁珠富集mRNA,加入片段化缓冲液(fragmentation buffer)将mRNA进行随机打断,以mRNA为模板,用六碱基随机引物合成第一条cDNA链;加入缓冲液、dNTPs、RNase H和DNA polymerase Ⅰ合成第二条cDNA链;利用AMPure XP磁珠纯化双链cDNA,进行末端修复、加A尾并连接测序接头,用AMPure XP beads进行片段大小选择,通过PCR富集得到cDNA文库。文库构建完成后,分别使用Qubit2.0荧光计和Agilent 2100生物分析仪对文库浓度和插入片段大小进行检测,使用qPCR对文库的有效浓度进行准确定量,以保证文库质量。质检合格后,用NovaSeq 6000测序平台进行高通量测序,采用150 bp的双端测序读长。使用Trim Galore软件对测序结果中的接头和低质量读长进行过滤和清除,使用STAR软件把测序结果比对到人类hg38基因组。比对完成后用featureCount读取转录本的数目,并用edgeR软件包进行标准化,标准化方法为logCPM(counts per million reads)。使用Hiplot Pro平台(https://hiplot.com.cn/)对应工具绘制转录差异基因热图,并进行基因本体论(Gene Ontology,GO)功能分析和京都基因和基因组数据库(Kyoto Encyclopedia of Genes and Genomes,KEGG)富集分析。
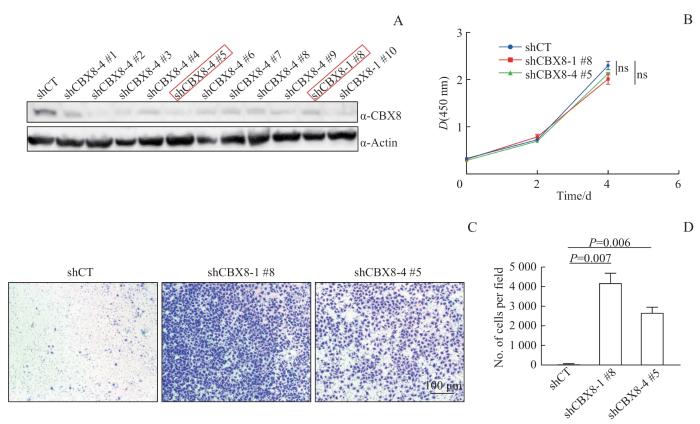
Note:A. Western blotting analysis demonstrating the expression levels of CBX8 after its stable knockdown in DU145 cell line. Selected cell lines for subsequent experiments are marked with red box. B. Effects of CBX8 knockdown on the proliferation of DU145 cell line were evaluated via CCK8 assay. C. The impact of CBX8 knockdown on the invasive potential of DU145 cell line was depicted by the Transwell invasion assay. Scale bar=100 μm. D. Statistical analysis derived from the aforementioned Transwell invasion assay results. ns—no statistical significance.
Fig 2
Effects of knocking down of CBX8 on the proliferation and invasion ability of prostate cancer cells
2.3 敲低 CBX8 后DU145细胞基因表达谱的改变及其差异分析
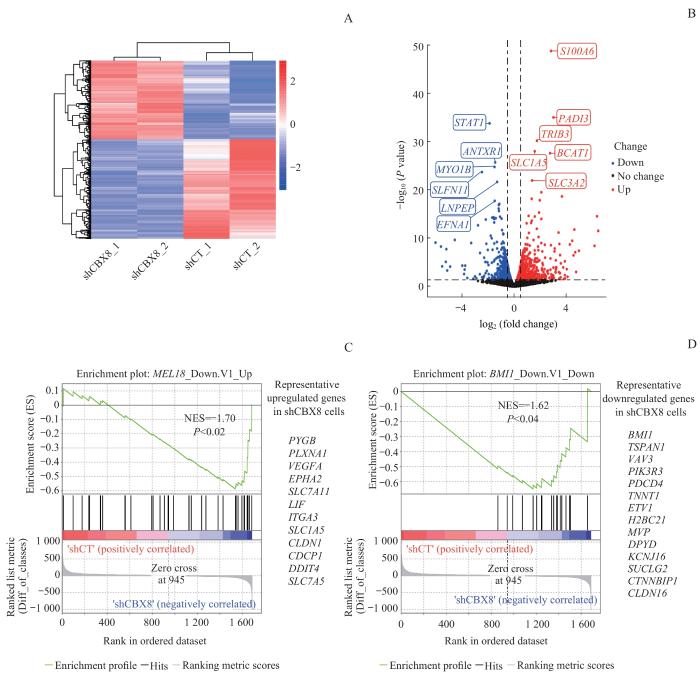
为了研究CBX8调控前列腺癌的作用机制,利用RNA-seq探究敲低CBX8后DU145细胞基因表达谱的变化。敲低CBX8导致750个基因上调,951个基因下调(图3A)。火山图(volcano plot)显示了表达上调和下调最为明显的基因(图3B),其中上调最明显的基因为蛋白S100A6(protein S100A6)、肽基精氨酸脱亚胺酶Ⅲ型(protein-arginine deiminase type-3,PADI3)、tribble同源蛋白3(tribbles homolog 3,TRIB3)、支链氨基酸转氨酶1(branched-chain-amino-acid aminotransferase 1,BCAT1)、溶质载体家族1成员5(solute carrier family 1 member 5,SLC1A5)、溶质载体家族3成员2(solute carrier family 3 member 2,SLC3A2);下调最明显的基因为信号转导和转录激活因子1(signal transducer and activator of transcription 1,STAT1)、炭疽毒素受体1(anthrax toxin receptor 1,ANTXR1)、肌球蛋白IB(myosin IB,MYO1B)、schlafen家族成员11(schlafen family member 11,SLFN11)、亮氨酰和胱氨酰氨肽酶(leucyl and cystinyl aminopeptidase,LNPEP)和肝配蛋白A1(ephrin-A1,EFNA1)。值得注意的是,上调的支链氨基酸代谢相关酶BCAT1与肺癌、肝癌等肿瘤的转移高度相关[23-24],一定程度上暗示敲低CBX8导致的DU145细胞侵袭能力升高可能是BCAT1介导的。另一方面,GSEA分析显示,敲低CBX8后上调的基因富集了敲低PRC1复合体另一核心组分多梳蛋白家族RING指蛋白2(polycomb group RING finger protein 2,PCGF2,又称MEL18)上调的基因(图3C),如糖原磷酸化酶脑型(glycogen phosphorylase brain form,PYGB)、丛蛋白A1(plexin-A1,PLXNA1)等;同样地,敲低CBX8后下调的基因也富集了敲低多梳蛋白家族RING指蛋白4(polycomb group RING finger protein 4,PCGF4,又称BMI1)下调的基因(图3D),如BMI1、四跨膜蛋白1(tetraspanin-1,TSPAN1)等。以上结果表明作为PRC1亚基之一的CBX8,其影响基因转录的作用机制很大程度是依赖于PRC1复合体功能。
Note: A. The heatmap of the differentially expressed genes after CBX8 knocking down in DU145 cell line. Color bar means log fold change. B. Volcano plot of differentially expressed genes after CBX8 knocking down in DU145 cell line. C. GSEA showed that genes differentially expressed after CBX8 knocking down were enriched in gene sets of upregulated genes when knocking down PCGF2 (MEL18). D. GSEA showed that genes differentially expressed after CBX8 knocking down were enriched in gene sets of downregulated genes when knocking down PCGF4 (BMI1). Gene sets in Fig 3C and D were obtained from MSigDB (The Molecular Signatures Database). NES—normalized enrichment score.
Fig 3
Gene expression profiling and GSEA of CBX8 knocking down prostate cancer cells
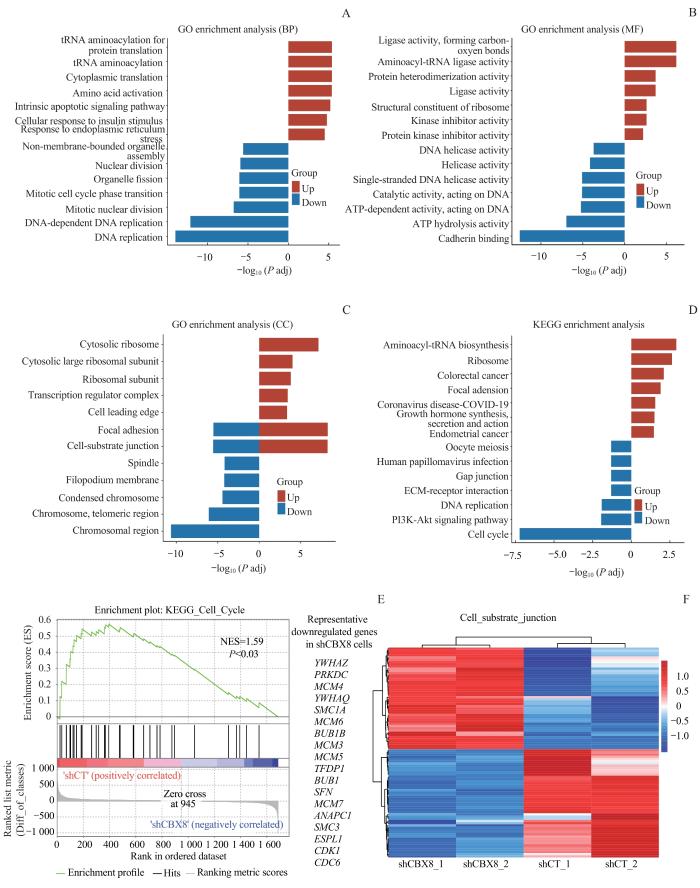
Note:A. GO biological process (BP) analysis of differentially expressed genes in CBX8 knocking down DU145 cell line. B. GO molecular function (MF) analysis of differentially expressed genes in CBX8 knocking down DU145 cell line. C. GO cell component (CC) analysis of differentially expressed genes in CBX8 knocking down DU145 cell line. D. KEGG analysis of differentially expressed genes in CBX8 knocking down DU145 cell line. P adj—adjusted P value. E. GSEA showed that genes differentially expressed after CBX8 knocking down were enriched in gene sets related to cell cycle. NES—normalized enrichment score. F. Heat map of genes differentially expressed in cell-substrate junction gene set after CBX8 knocking down.
Fig 4
Functional annotation and enrichment analysis of differentially expressed genes in CBX8 knocking down prostate cancer cells
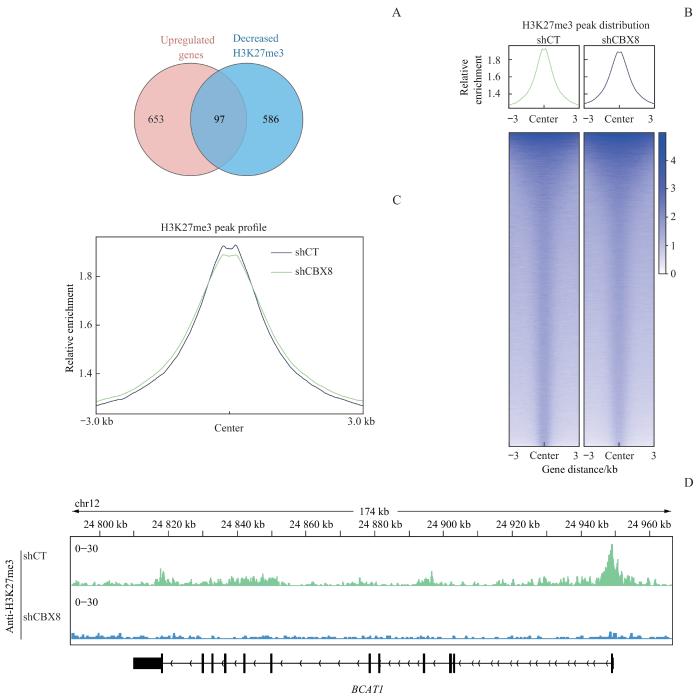
Note:A. The Venn diagram of unique and shared genes between upregulated genes and genes with decreased H3K27me3 peaks after CBX8 knocking down. B. Peak intensity analysis of H3K27me3 in shCT and shCBX8-treated DU145 cells. C. Heatmap analysis of H3K27me3 enrichment in shCT and shCBX8-treated DU145 cells. D. Overview of H3K27me3 occupancy at the BCAT1 promoter region in shCT and shCBX8-treated DU145 cells.
Fig 5
Effect of CBX8 knocking down on H3K27me3 modification in prostate cancer cells
YANG Wanli, SONG Juan and LAO Yimin conducted experiments and analyzed the collected data. YANG Wanli and LAO Yimin contributed to the idea of the study and wrote the paper. LI Bing supervised the whole project development, paper writing and editing. All the authors have read the last version of paper and agreed for the submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
PIUNTI A, SHILATIFARD A. The roles of Polycomb repressive complexes in mammalian development and cancer[J]. Nat Rev Mol Cell Biol, 2021, 22(5): 326-345.
SU W J, HAN H H, WANG Y, et al. The polycomb repressor complex 1 drives double-negative prostate cancer metastasis by coordinating stemness and immune suppression[J]. Cancer Cell, 2019, 36(2): 139-155.e10.
JUNG J, BUISMAN S C, WEERSING E, et al. CBX7 induces self-renewal of human normal and malignant hematopoietic stem and progenitor cells by canonical and non-canonical interactions[J]. Cell Rep, 2019, 26(7): 1906-1918.e8.
KLAUKE K, RADULOVIĆ V, BROEKHUIS M, et al. Polycomb Cbx family members mediate the balance between haematopoietic stem cell self-renewal and differentiation[J]. Nat Cell Biol, 2013, 15(4): 353-362.
DEL GAUDIO N, DI COSTANZO A, LIU N Q, et al. CBX2 shapes chromatin accessibility promoting AML via p38 MAPK signaling pathway[J]. Mol Cancer, 2022, 21(1): 125.
HU F F, CHEN H, DUAN Y, et al. CBX2 and EZH2 cooperatively promote the growth and metastasis of lung adenocarcinoma[J]. Mol Ther Nucleic Acids, 2021, 27: 670-684.
WANG X, QIN G, LIANG X T, et al. Targeting the CK1α/CBX4 axis for metastasis in osteosarcoma[J]. Nat Commun, 2020, 11(1): 1141.
ZHANG C Z, CHEN S L, WANG C H, et al. CBX8 exhibits oncogenic activity via AKT/β-catenin activation in hepatocellular carcinoma[J]. Cancer Res, 2018, 78(1): 51-63.
MENG Q C, LI L, WANG L P. High CBX8 expression leads to poor prognosis in laryngeal squamous cell carcinoma by inducing EMT by activating the Wnt/β-catenin signaling pathway[J]. Front Oncol, 2022, 12: 881262.
DENG H L, GUAN X W, GONG L C, et al. CBX6 is negatively regulated by EZH2 and plays a potential tumor suppressor role in breast cancer[J]. Sci Rep, 2019, 9(1): 197.
HUANG Z N, YAN Y L, ZHU Z, et al. CBX7 suppresses urinary bladder cancer progression via modulating AKR1B10-ERK signaling[J]. Cell Death Dis, 2021, 12(6): 537.
BERNARD D, MARTINEZ-LEAL J F, RIZZO S, et al. CBX7 controls the growth of normal and tumor-derived prostate cells by repressing the Ink4a/Arf locus[J]. Oncogene, 2005, 24(36): 5543-5551.
ZHANG Y, KANG M, ZHANG B, et al. m6A modification-mediated CBX8 induction regulates stemness and chemosensitivity of colon cancer via upregulation of LGR5[J]. Mol Cancer, 2019, 18(1): 185.
CONNELLY K E, WEAVER T M, ALPSOY A, et al. Engagement of DNA and H3K27me3 by the CBX8 chromodomain drives chromatin association[J]. Nucleic Acids Res, 2019, 47(5): 2289-2305.
CERAMI E, GAO J J, DOGRUSOZ U, et al. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data[J]. Cancer Discov, 2012, 2(5): 401-404.
SUBRAMANIAN A, TAMAYO P, MOOTHA V K, et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles[J]. Proc Natl Acad Sci USA, 2005, 102(43): 15545-15550.
MAO L, CHEN J, LU X, et al. Proteomic analysis of lung cancer cells reveals a critical role of BCAT1 in cancer cell metastasis[J]. Theranostics, 2021, 11(19): 9705-9720.
TANG J J, WANG G, ZHANG M F, et al. Paradoxical role of CBX8 in proliferation and metastasis of colorectal cancer[J]. Oncotarget, 2014, 5(21): 10778-10790.
SHANG S, YANG Y W, CHEN F, et al. TRIB3 reduces CD8+ T cell infiltration and induces immune evasion by repressing the STAT1-CXCL10 axis in colorectal cancer[J]. Sci Transl Med, 2022, 14(626): eabf0992.
JIANG Q, LI J, WANG J Y, et al. Inhibition of CDKL3 downregulates STAT1 thus suppressing prostate cancer development[J]. Cell Death Dis, 2023, 14(3): 189.
HATZIIEREMIA S, MOHAMMED Z, MCCALL P, et al. Loss of signal transducer and activator of transcription 1 is associated with prostate cancer recurrence[J]. Mol Carcinog, 2016, 55(11): 1667-1677.
KACHROO P, LEE M H, ZHANG L, et al. IL-27 inhibits epithelial-mesenchymal transition and angiogenic factor production in a STAT1-dominant pathway in human non-small cell lung cancer[J]. J Exp Clin Cancer Res, 2013, 32(1): 97.
WANG T, HAN S L, DU G. S100A6 represses Calu-6 lung cancer cells growth via inhibiting cell proliferation, migration, invasion and enhancing apoptosis[J]. Cell Biochem Funct, 2021, 39(6): 771-779.
MACRAE T A, FOTHERGILL-ROBINSON J, RAMALHO-SANTOS M. Regulation, functions and transmission of bivalent chromatin during mammalian development[J]. Nat Rev Mol Cell Biol, 2023, 24(1): 6-26.
TERRANOVA C J, TANG M, MAITITUOHETI M, et al. Reprogramming of bivalent chromatin states in NRAS mutant melanoma suggests PRC2 inhibition as a therapeutic strategy[J]. Cell Rep, 2021, 36(3): 109410.
CHAN H L, BECKEDORFF F, ZHANG Y S, et al. Polycomb complexes associate with enhancers and promote oncogenic transcriptional programs in cancer through multiple mechanisms[J]. Nat Commun, 2018, 9(1): 3377.
... 应用基因集富集分析(Gene Set Enrichment Analysis,GSEA)软件[21]对RNA-seq结果进行基因富集分析和前沿分析(leading edge analysis),参考基因集数据选取自分子特征数据库(Molecular Signatures Database,MSigDB)[22].基因排名方式(metric for ranking genes)选择Diff_of_classes,其余选项为默认.代表性变化基因由软件根据基因排名选出. ...
3
... 为了研究CBX8调控前列腺癌的作用机制,利用RNA-seq探究敲低CBX8后DU145细胞基因表达谱的变化.敲低CBX8导致750个基因上调,951个基因下调(图3A).火山图(volcano plot)显示了表达上调和下调最为明显的基因(图3B),其中上调最明显的基因为蛋白S100A6(protein S100A6)、肽基精氨酸脱亚胺酶Ⅲ型(protein-arginine deiminase type-3,PADI3)、tribble同源蛋白3(tribbles homolog 3,TRIB3)、支链氨基酸转氨酶1(branched-chain-amino-acid aminotransferase 1,BCAT1)、溶质载体家族1成员5(solute carrier family 1 member 5,SLC1A5)、溶质载体家族3成员2(solute carrier family 3 member 2,SLC3A2);下调最明显的基因为信号转导和转录激活因子1(signal transducer and activator of transcription 1,STAT1)、炭疽毒素受体1(anthrax toxin receptor 1,ANTXR1)、肌球蛋白IB(myosin IB,MYO1B)、schlafen家族成员11(schlafen family member 11,SLFN11)、亮氨酰和胱氨酰氨肽酶(leucyl and cystinyl aminopeptidase,LNPEP)和肝配蛋白A1(ephrin-A1,EFNA1).值得注意的是,上调的支链氨基酸代谢相关酶BCAT1与肺癌、肝癌等肿瘤的转移高度相关[23-24],一定程度上暗示敲低CBX8导致的DU145细胞侵袭能力升高可能是BCAT1介导的.另一方面,GSEA分析显示,敲低CBX8后上调的基因富集了敲低PRC1复合体另一核心组分多梳蛋白家族RING指蛋白2(polycomb group RING finger protein 2,PCGF2,又称MEL18)上调的基因(图3C),如糖原磷酸化酶脑型(glycogen phosphorylase brain form,PYGB)、丛蛋白A1(plexin-A1,PLXNA1)等;同样地,敲低CBX8后下调的基因也富集了敲低多梳蛋白家族RING指蛋白4(polycomb group RING finger protein 4,PCGF4,又称BMI1)下调的基因(图3D),如BMI1、四跨膜蛋白1(tetraspanin-1,TSPAN1)等.以上结果表明作为PRC1亚基之一的CBX8,其影响基因转录的作用机制很大程度是依赖于PRC1复合体功能. ...
... 为了研究CBX8调控前列腺癌的作用机制,利用RNA-seq探究敲低CBX8后DU145细胞基因表达谱的变化.敲低CBX8导致750个基因上调,951个基因下调(图3A).火山图(volcano plot)显示了表达上调和下调最为明显的基因(图3B),其中上调最明显的基因为蛋白S100A6(protein S100A6)、肽基精氨酸脱亚胺酶Ⅲ型(protein-arginine deiminase type-3,PADI3)、tribble同源蛋白3(tribbles homolog 3,TRIB3)、支链氨基酸转氨酶1(branched-chain-amino-acid aminotransferase 1,BCAT1)、溶质载体家族1成员5(solute carrier family 1 member 5,SLC1A5)、溶质载体家族3成员2(solute carrier family 3 member 2,SLC3A2);下调最明显的基因为信号转导和转录激活因子1(signal transducer and activator of transcription 1,STAT1)、炭疽毒素受体1(anthrax toxin receptor 1,ANTXR1)、肌球蛋白IB(myosin IB,MYO1B)、schlafen家族成员11(schlafen family member 11,SLFN11)、亮氨酰和胱氨酰氨肽酶(leucyl and cystinyl aminopeptidase,LNPEP)和肝配蛋白A1(ephrin-A1,EFNA1).值得注意的是,上调的支链氨基酸代谢相关酶BCAT1与肺癌、肝癌等肿瘤的转移高度相关[23-24],一定程度上暗示敲低CBX8导致的DU145细胞侵袭能力升高可能是BCAT1介导的.另一方面,GSEA分析显示,敲低CBX8后上调的基因富集了敲低PRC1复合体另一核心组分多梳蛋白家族RING指蛋白2(polycomb group RING finger protein 2,PCGF2,又称MEL18)上调的基因(图3C),如糖原磷酸化酶脑型(glycogen phosphorylase brain form,PYGB)、丛蛋白A1(plexin-A1,PLXNA1)等;同样地,敲低CBX8后下调的基因也富集了敲低多梳蛋白家族RING指蛋白4(polycomb group RING finger protein 4,PCGF4,又称BMI1)下调的基因(图3D),如BMI1、四跨膜蛋白1(tetraspanin-1,TSPAN1)等.以上结果表明作为PRC1亚基之一的CBX8,其影响基因转录的作用机制很大程度是依赖于PRC1复合体功能. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}