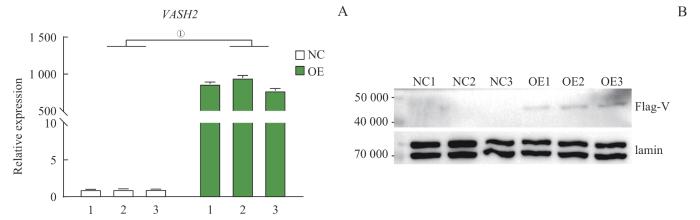
目的·探索血管抑制蛋白2(vasohibin-2,VASH2)在三阴性乳腺癌(triple-negative breast cancer,TNBC)中的表达及VASH2通过调控基因表达和可变剪接在TNBC发生发展中的作用机制。方法·通过基因型-组织表达数据库(Genotype-Tissue Expression,GTEx)联合癌症基因组图谱数据库(The Cancer Genome Atlas,TCGA)比较VASH2在TNBC中与正常乳腺组织中的表达差异,并分析VASH2在TNBC中的甲基化水平。在TNBC细胞系MDA-MB-231中对VASH2过表达,并进行转录组测序,分析受VASH2所调控的差异表达基因及可变剪接基因。结果·VASH2在TNBC组织中与正常组织及其他乳腺癌分型比较表达水平显著提高。VASH2基因的低甲基化可能是VASH2在TNBC 中表达上调的原因之一。VASH2过表达后引起81个基因显著差异表达,其中上调的基因23个,下调的基因58个。VASH2过表达后可变剪接水平发生显著变化的基因主要富集在细胞周期、p53信号通路上。结论·VASH2可能通过调控TNBC中基因可变剪接促进其发生和发展。
关键词:血管抑制蛋白2
;
三阴性乳腺癌
;
可变剪接
;
p53信号通路
;
细胞周期
Abstract
Objective ·To explore the role of vasohibin-2 (VASH2) in the regulation of proliferation and metastasis of triple-negative breast cancer (TNBC) cells, and explore the mechanism of VASH2 in the occurrence and development of TNBC through regulation of gene expression and alternative splicing. Methods ·TCGA-GTEx was used to analyze the expression of VASH2 in TNBC. VASH2 methylation levels in TNBC were also analyzed. VASH2 was overexpressed in the MDA-MB-231 human TNBC cell line and transcriptome sequencing was performed. Differentially expressed genes and alternatively spliced genes regulated by VASH2 were analyzed to explore the mechanism of action of VASH2 in TNBC. Results ·VASH2 was significantly overexpressed in TNBC compared to the normal tissues. Hypomethylation of the VASH2 gene was implicated in the upregulation of VASH2 expression in TNBC. Overexpression of VASH2 caused significant differential expression of 81 genes, of which 23 genes were up-regulated and 58 genes were down-regulated. Genes with significantly altered alternative splicing levels due to VASH2 overexpressed were enriched in cell cycle and p53 signaling pathways. Conclusion ·VASH2 regulates the alternative splicing of TNBC oncogenes and promotes TNBC occurrence and development.
Keywords:vasohibin-2 (VASH2)
;
triple-negative breast cancer (TNBC)
;
alternative splicing
;
p53 signaling pathway
;
cell cycle
WANG Wei, WANG Hongli, ALIBIYATI·i Ain, YILIYAER· Rousu, AYI NUER, YANG Liang. Function of vasohibin-2 and the mechanism of alternative splicing in triple-negative breast cancer. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2024, 44(12): 1526-1535 doi:10.3969/j.issn.1674-8115.2024.12.005
三阴性乳腺癌(triple-negative breast cancer,TNBC)是一种肿瘤异质性高、复发率高、进展快、预后差、治疗抵抗相关的特殊乳腺癌(breast cancer,BC)亚型[1-3]。有研究显示,血管生成在BC发生、发展、侵袭以及转移等过程中发挥着至关重要的作用[4]。新生血管的营养支持为肿瘤组织的持续生长提供可能,从而促进肿瘤细胞的分裂增殖,以及肿瘤细胞向远处的浸润和转移[5]。血管抑制蛋白2(vasohibin-2,VASH2)是一种新发现的具有促进新生血管形成和进展作用的血管调节因子,其主要表达在肿瘤细胞和单核细胞血管新生区[6]。其在TNBC中呈现高表达,与TNBC更容易发生内脏转移密切相关[7]。已证实,VASH2与多种人类肿瘤的增殖、转移和侵袭相关[8-10],能通过激活TGF-β信号通路,参与上皮间质转化过程的调控进而增强卵巢癌和BC细胞的迁移与侵袭能力[11-12],但其在TNBC中侵袭和转移的分子机制尚不清楚。本研究即对VASH2在TNBC中的表达及其作用机制进行了探索。
1 资料与方法
1.1 一般资料
由于癌症基因组图谱数据库(The Cancer Genome Atlas,TCGA)中BC的癌旁正常组织样本较少,因此从UCSC xena网站下载基因型-组织表达数据库(Genotype-Tissue Expression,GTEx)与TCGA的FPKM格式的测序数据,筛选出TNBC样本。使用R及perl软件对选取的TCGA转录组数据进行预处理,剔除不合格样本后,再将数据进行合并、汇总、注释。另外,从UCSC xena下载Illumina Human Methylation450芯片测序数据,用于甲基化数据分析。
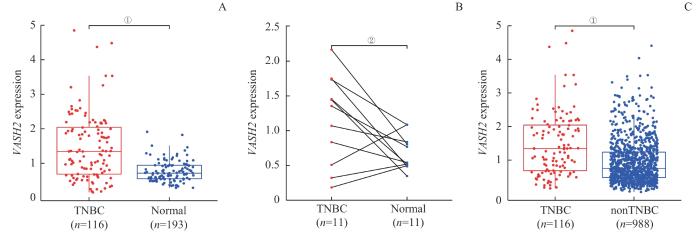
Note: A. Differential expression between TNBC and normal breast tissues. B. Differential expression between TNBC and paired cancer-adjacent tissues. C. Comparative analysis between TNBC and nonTNBC. ①P<0.001, ②P=0.035.
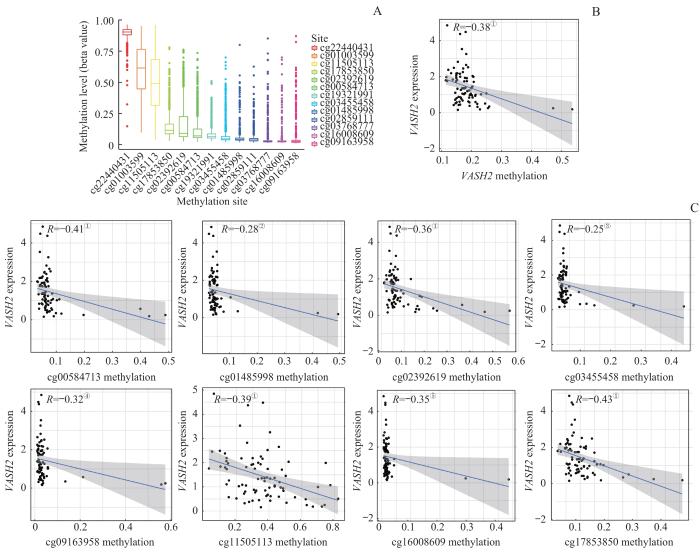
Note: A. Methylation site correlation analysis in the VASH2 promoter. B. Methylation levels corresponded to VASH2 expression. C. Correlation analysis between the degree of methylation sites and the expression of VASH2. ①P<0.001, ②P=0.011, ③P=0.029, ④P=0.004, ⑤P=0.002.
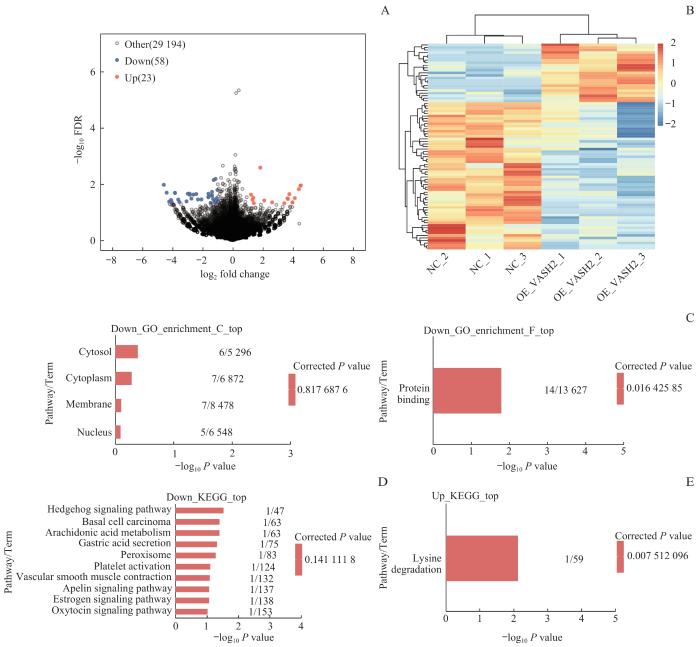
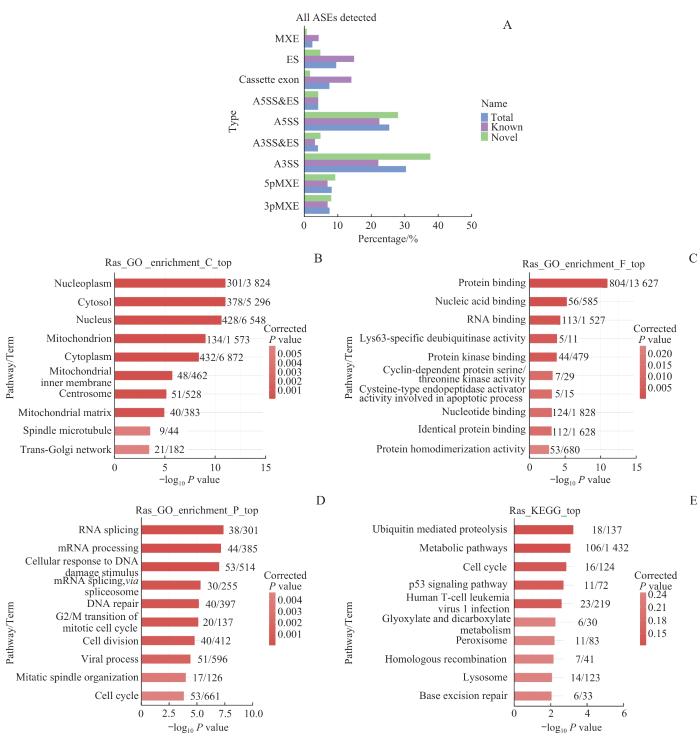
Note: A. Volcano map based on differentially expressed genes (DEGs). B. Heat map of expression levels of DEGs. C. GO function enrichment bar chart. D/E. Bar charts showing KEGG pathway enrichment of downregulated DEGs (D) and upregulated DEGs (E).
Fig 4
RNA-seq analysis of VASH2-regulated transcriptome
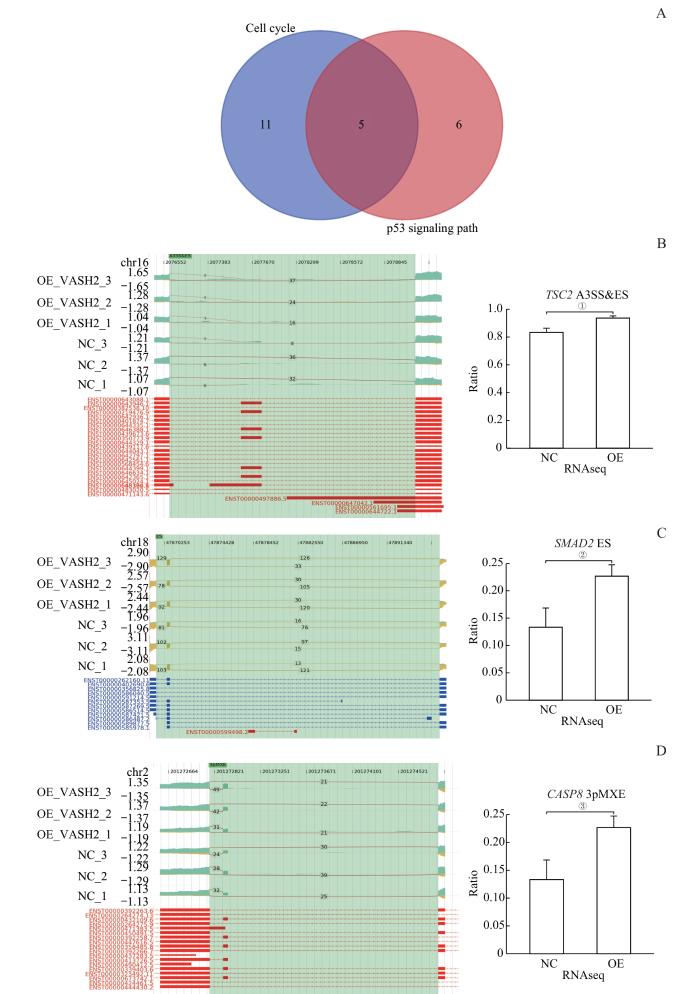
Note: A. Wayne diagram of the gene set with significantly altered AS expression levels in the cell cycle and p53 signaling pathway. B. ASE of the VASH2-regulated gene TSC2 and IGV-sashimi plot showing A3SS & ES events. C.ASE of the VASH2-regulated gene SMAD2 and IGV-sashimi plot showing ES events. D. ASE of the VASH2-regulated gene CASP8. ①P=0.006, ②P=0.017, ③P=0.027.
Fig 6
Apoptosis-enriched alternative splicing genes
YANG Liang and WANG Wei conducted the experiment, acquired and analyzed the data, and drafted the manuscript. WANG Hongli and ALIBIYATI·Aini participated in manuscript modification. YILIYAER·Rousu designed and guided the study and revised the manuscript. AYINUER participated in study guidance. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
ZONG Y, PEGRAM M. Research advances and new challenges in overcoming triple-negative breast cancer[J]. Cancer Drug Resist, 2021, 4(3): 517-542.
MERIKHIAN P, EISAVAND M R, FARAHMAND L. Triple-negative breast cancer: understanding Wnt signaling in drug resistance[J]. Cancer Cell Int, 2021, 21(1): 419.
BURSTEIN H J, CURIGLIANO G, THÜRLIMANN B, et al. Customizing local and systemic therapies for women with early breast cancer: the St. Gallen International Consensus Guidelines for treatment of early breast cancer 2021[J]. Ann Oncol. 2021,32(10):1216-1235.
TU J J, FANG Y L, HAN D F, et al. Activation of nuclear factor-κB in the angiogenesis of glioma: insights into the associated molecular mechanisms and targeted therapies[J]. Cell Prolif, 2021, 54(2): e12929.
TAN X B, LIAO Z F, ZOU S Y, et al. VASH2 promotes cell proliferation and resistance to doxorubicin in non-small cell lung cancer via AKT signaling[J]. Oncol Res, 2020, 28(1): 3-11.
NINOMIYA Y, OZAWA S, OGUMA J, et al. Expression of vasohibin-1 and-2 predicts poor prognosis among patients with squamous cell carcinoma of the esophagus[J]. Oncol Lett, 2018, 16(4): 5265-5274.
YAMAMOTO M, OZAWA S, NINOMIYA Y, et al. Plasma vasohibin-1 and vasohibin-2 are useful biomarkers in patients with esophageal squamous cell carcinoma[J]. Esophagus, 2020, 17(3): 289-297.
KITAHARA S, SUZUKI Y, MORISHIMA M, et al. Vasohibin-2 modulates tumor onset in the gastrointestinal tract by normalizing tumor angiogenesis[J]. Mol Cancer, 2014, 13: 99.
NORITA R, SUZUKI Y, FURUTANI Y, et al. Vasohibin-2 is required for epithelial-mesenchymal transition of ovarian cancer cells by modulating transforming growth factor-β signaling[J]. Cancer Sci, 2017, 108(3): 419-426.
TU M, LI Z J, LIU X, et al. Vasohibin 2 promotes epithelial-mesenchymal transition in human breast cancer via activation of transforming growth factor β 1 and hypoxia dependent repression of GATA-binding factor 3[J]. Cancer Lett, 2017, 388: 187-197.
MA H R, CAO L, WANG F, et al. Filamin B extensively regulates transcription and alternative splicing, and is associated with apoptosis in HeLa cells[J]. Oncol Rep, 2020, 43(5): 1536-1546.
YISA S B, FEI W M, YAXUN W M, et al. ATP5A1 participates in transcriptional and posttranscriptional regulation of cancer-associated genes by modulating their expression and alternative splicing profiles in HeLa cells[J]. Technol Cancer Res Treat, 2021, 20: 15330338211039126.
SHEN X X, JIN L, ZHAO Q, et al. Expression of vasohibin-2 and its effect on prognosis of breast cancer[J]. Anhui Medical Journal, 2021, 42(2): 144-148.
GAO Y, ZHANG W Z, LIU C W, et al. MiR-200 affects tamoxifen resistance in breast cancer cells through regulation of MYB[J]. Sci Rep, 2019, 9(1): 18844.
ZENG X, QU X J, ZHAO C Y, et al. FEN1 mediates miR-200a methylation and promotes breast cancer cell growth via MET and EGFR signaling[J]. FASEB J, 2019, 33(10): 10717-10730.
ANSARI J, SHACKELFORD R E, EL-OSTA H. Epigenetics in non-small cell lung cancer: from basics to therapeutics[J]. Transl Lung Cancer Res, 2016, 5(2): 155-171.
LI R, YANG Y E, YIN Y H, et al. Methylation and transcriptome analysis reveal lung adenocarcinoma-specific diagnostic biomarkers[J]. J Transl Med, 2019, 17(1): 324.
CHEN Y C, LIU X R, LI Y K, et al. Lung cancer therapy targeting histone methylation: opportunities and challenges[J]. Comput Struct Biotechnol J, 2018, 16: 211-223.
WANG B, YANG L, ZHAO Q, et al. Vasohibin 2 as a potential predictor of aggressive behavior of triple-negative breast cancer[J]. Am J Transl Res, 2017, 9(6): 2911-2919.
VAN DER LAAN S, LÉVÊQUE M F, MARCELLIN G, et al. Evolutionary divergence of enzymatic mechanisms for tubulin detyrosination[J]. Cell Rep, 2019, 29(12): 4159-4171.e6.
NIEUWENHUIS J, ADAMOPOULOS A, BLEIJERVELD O B, et al. Vasohibins encode tubulin detyrosinating activity[J]. Science, 2017, 358(6369): 1453-1456.
JOERGER A C, FERSHT A R. The p53 pathway: origins, inactivation in cancer, and emerging therapeutic approaches[J]. Annu Rev Biochem, 2016, 85: 375-404.
ZHANG H, ZHANG X, LI X, et al. Effect of CCNB1 silencing on cell cycle, senescence, and apoptosis through the p53 signaling pathway in pancreatic cancer[J]. J Cell Physiol, 2018, 234(1): 619-631.
MISSAOUI N, LANDOLSI H, MESTIRI S, et al. Immunohistochemical analysis of c-erbB-2, Bcl-2, p53, p21(WAF1/Cip1), p63 and Ki-67 expression in hydatidiformmoles[J]. Pathol Res Pract,2019,215(3):446-452.
LO W, PARKHURST M, ROBBINS P F, et al. Immunologic recognition of a shared p53 mutated neoantigen in a patient with metastatic colorectal cancer[J]. Cancer Immunol Res,2019,7(4):534-543.
SCHAEFER I M, HORNICK J L, SHOLL L M, et al. Abnormal p53 and p16 staining patterns distinguish uterine leiomyosarcoma from inflammatory myofibroblastic tumour[J]. Histopathology, 2017, 70(7): 1138-1146.
HUANG X, SHAO D, WU H W, et al. Genomic profiling comparison of germline BRCA and non-BRCA carriers reveals CCNE1 amplification as a risk factor for non-BRCA carriers in patients with triple-negative breast cancer[J]. Front Oncol, 2020, 10: 583314.
YANG R, CHEN J X. Effects of circular RNA hsa_circ_0058514 on the development and progression of triple-negative breast cancer[J]. China Oncology, 2019, 29(1): 9-18.
LIU Z, FU Q S, WANG Y, et al. Synergy between vinorelbine and afatinib in the inhibition of non-small cell lung cancer progression by EGFR and p53 signaling pathways[J]. Biomed Pharmacother, 2021, 134: 111144.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}