Objective ·To investigate the effects of multiple sevoflurane exposures on the proliferation and differentiation of primary oligodendrocytes. Methods ·Oligodendrocyte precursor cells (OPCs) were extracted from the cortex of rats on the day of birth and cultured in vitro. The cells were divided into control and sevoflurane groups. To simulate the clinical situation of sevoflurane exposure, cells in the sevoflurane group were exposed to 3% sevoflurane for 3 consecutive days, 2 h for each time. After the OPCs were differentiated and matured, immunofluorescence staining and Western blotting were used to detect the expression of myelin basic protein (MBP) and the myelin-associated glycoprotein (MAG). Cell proliferation assays (BrdU and Ki67) and a cell viability assay (CCK8) were used to detect the effects of sevoflurane on the proliferation ability of OPCs and the survival rate of oligodendrocytes. Western blotting was used to detect the protein content of caspase-3. Lentiviral transfection technology was used to overexpress YTH N6-methyladenosine RNA binding protein F1 (YTHDF1) in OPCs, and then CCK8 was used to detect cell proliferation and survival. Results ·Immunofluorescence results showed that multiple sevoflurane exposures led to a decrease in the number of primary oligodendrocytes expressing mature myelin surface markers (MBP, MAG); Western blotting results showed that these exposures led to upregulation of caspase-3 expression in primary OPCs; CCK8 results showed that the survival rate of primary OPCs decreased with the increase in the number of sevoflurane treatments; however, BrdU and Ki67 staining results showed that the proliferation ability of primary OPCs was enhanced after sevoflurane exposure. In addition, overexpression of YTHDF1 could partially improve the decreased survival rate of primary OPCs caused by multiple sevoflurane exposures (all P<0.05). Conclusion ·Multiple sevoflurane exposures impair the myelinating ability and survival rate of primary oligodendrocytes, manifested by apoptosis of some primary OPCs. In contrast, sevoflurane exposure compensatorily increases the proliferation ability of surviving primary OPCs.
SHI Lingling, CHENG Yanyong, ZHANG Lei. Effects of sevoflurane exposure on proliferation and differentiation of primary oligodendrocytes. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2024, 44(9): 1115-1123 doi:10.3969/j.issn.1674-8115.2024.09.006
婴幼儿时期,人类大脑快速发育,对作用于神经系统的药物特别敏感。由于全身麻醉药物可以通过血脑屏障直接作用于脑,因此全身麻醉药物对发育脑的影响是值得重点关注的公共卫生问题之一。七氟烷是目前临床上婴幼儿患者最常用的吸入全身麻醉药物。梅奥儿童麻醉安全性研究(Mayo Anesthesia Safety in Kids,MASK)提出多次接触全身麻醉会影响儿童的精细运动能力,且在二次分析后提出部分儿童对多次接触的麻醉药物更为敏感[1]。2020年的一项研究[2]对13 433名儿童进行详细的神经发育评估。该研究涵盖了46项不同的神经发育指标,结果显示,频繁接触七氟烷的儿童在运动功能、手部灵活性以及社交能力方面的表现普遍较差。动物研究发现出生早期长时间接受七氟烷麻醉或暴露于高浓度的七氟烷会导致发育脑的神经毒性,例如焦虑相关行为[3-4],并增加成年期认知功能障碍的可能性[5]。这些发现表明七氟烷麻醉对神经发育具有潜在风险[6]。基础研究更多地关注了七氟烷对神经元的影响,例如七氟烷暴露可诱导神经元凋亡、破坏突触可塑性并损害认知功能[7-11],而非神经元细胞在中枢神经系统中也起到了举足轻重的作用。
出生当日(postnatal day 0,P0)的SD大鼠(性别不限),无特定病原体(specific pathogen free,SPF)级,由浙江维通利华实验动物技术有限公司提供。实验动物生产许可证号:SCXK(浙)2024-0001。动物实验于上海交通大学医学院附属第九人民医院中心实验室进行。实验动物使用许可证号:SYXK(沪)2020-0025。
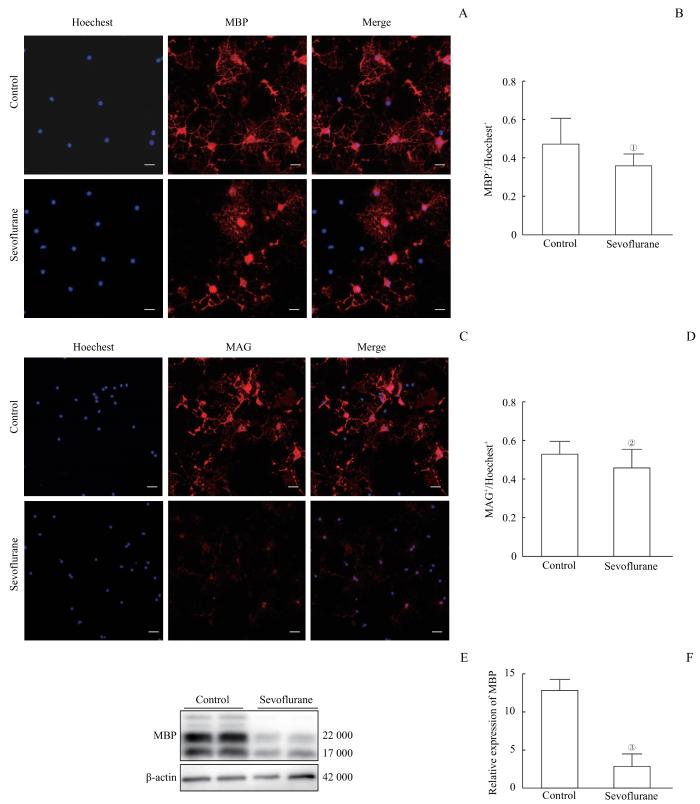
Note: A. Immunofluorescence staining showed the expression of MBP in oligodendrocytes. The scale bar is 100 μm. B. The number of Hoechest-positive cells normalized the number of MBP-positive cells. Multiple sevoflurane exposures significantly reduced the number of MBP-expressing cells. C. Immunofluorescence staining showed the expression of MAG in oligodendrocytes. The scale bar is 100 μm. D. The number of MAG-positive cells was normalized by the number of Hoechest-positive cells. Multiple sevoflurane exposures significantly reduced the number of MAG-expressing cells. E. Detection of MBP expression levels in oligodendrocytes by Western blotting. F. Western blotting results showed that multiple sevoflurane exposures significantly reduced the expression of MBP protein levels. ①P=0.030, ②P=0.026, ③P=0.001, compared with the control group.
Fig 1
Effects of multiple sevoflurane exposures on the myelinating ability of oligodendrocytes
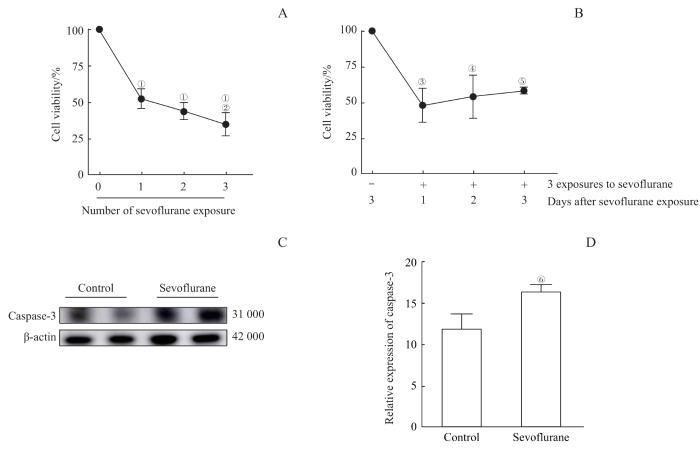
Note: A/B. CCK8 results suggested that sevoflurane had a killing effect on OPCs. A. 1, 2, and 3 treatments with sevoflurane all had a killing effect on OPCs. As the number of sevoflurane exposures increased, the survival rate of OPCs decreased. B. The survival rate of OPCs decreased significantly on day 1, 2, and 3 after sevoflurane exposure, but there was no significant difference between each detection day. C. Detection of caspase-3 expression levels in OPCs by Western blotting. D. Western blotting showed that multiple sevoflurane exposures increased the expression of caspase-3 protein levels. ①P=0.000, compared with the group with 0 sevoflurane exposure; ②P=0.020, compared with the group with 1 sevoflurane exposure; ③P=0.011, ④P=0.030, ⑤P=0.046, compared with the group without sevoflurane exposure; ⑥P=0.012, compared with the control group.
Fig 2
Toxic effects of multiple sevoflurane exposure on OPCs by inducing apoptosis
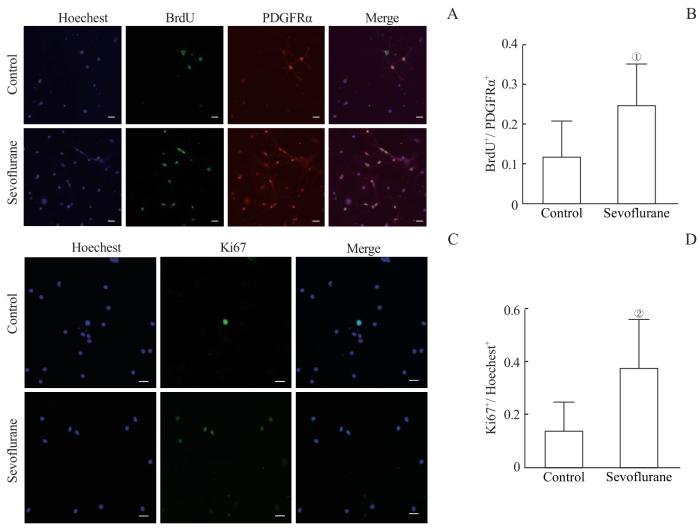
Note: A. Immunofluorescence staining showed the expression of BrdU in OPCs. The scale bar is 100 μm. B. The number of BrdU-positive cells was normalized to the number of PDGFRα positive cells. Multiple sevoflurane exposures increased the proliferation level of PDGFRα-positive OPCs. C. Immunofluorescence staining showed the expression of Ki67 in OPCs. The scale bar is 100 μm. D. The number of Ki67-positive cells was normalized to the number of Hoechest-positive cells. Multiple sevoflurane exposures increased the proliferation level of PDGFRα-positive OPCs. ①P=0.008, ②P=0.007, compared with the control group.
Fig 3
Effects of multiple sevoflurane exposure on the proliferation of OPCs
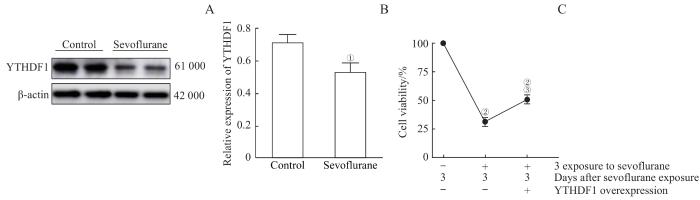
Note: A. Detection of YTHDF1 expression levels in OPCs by Western blotting. B. Western blotting results showed that multiple sevoflurane exposures reduced the expression level of YTHDF1 protein. C. CCK8 experiments indicated that sevoflurane exposures killed OPCs and significantly reduced cell survival rate. However, cells overexpressing YTHDF1 had a partial protective effect on the killing effect of sevoflurane. ①P=0.011, compared with the control group; ②P=0.000, compared with the group without sevoflurane exposure and with control virus infection; ③P=0.007, compared with the group with 3 sevoflurane exposures and control virus infection.
Fig 4
Overexpression of YTHDF1 rescued the damage of OPCs caused by multiple sevoflurane exposures
CHENG Yanyong and ZHANG Lei participated in the experimental design; SHI Lingling completed the experimental operation and data analysis; SHI Lingling, CHENG Yanyong, and ZHANG Lei participated in the writing and revision of the paper. All authors have read the final manuscript and approved for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interest.
WARNER D O, ZACCARIELLO M J, KATUSIC S K, et al. Neuropsychological and behavioral outcomes after exposure of young children to procedures requiring general anesthesia: the Mayo Anesthesia Safety in Kids (MASK) study[J]. Anesthesiology, 2018, 129(1): 89-105.
WALKDEN G J, GILL H, DAVIES N M, et al. Early childhood general anesthesia and neurodevelopmental outcomes in the Avon longitudinal study of parents and children birth cohort[J]. Anesthesiology, 2020, 133(5): 1007-1020.
RAPER J, DE BIASIO J C, MURPHY K L, et al. Persistent alteration in behavioural reactivity to a mild social stressor in rhesus monkeys repeatedly exposed to sevoflurane in infancy[J]. Br J Anaesth, 2018, 120(4): 761-767.
ROSADO-MENDEZ I M, NOGUCHI K K, CASTAÑEDA-MARTINEZ L, et al. Quantitative ultrasound and apoptotic death in the neonatal primate brain[J]. Neurobiol Dis, 2019, 127: 554-562.
DAI C L, LI H C, HU X, et al. Neonatal exposure to anesthesia leads to cognitive deficits in old age: prevention with intranasal administration of insulin in mice[J]. Neurotox Res, 2020, 38(2): 299-311.
XIE L H, LIU Y, HU Y H, et al. Neonatal sevoflurane exposure induces impulsive behavioral deficit through disrupting excitatory neurons in the medial prefrontal cortex in mice[J]. Transl Psychiatry, 2020, 10(1): 202.
JIANG J, LV X, WU X Y, et al. Downregulation of circulating insulin-like growth factor 1 contributes to memory impairment in aged mice after sevoflurane anesthesia[J]. Behav Pharmacol, 2017, 28(2 and 3-Spec Issue): 238-243.
SHEN Y S, ZHOU T, LIU X B, et al. Sevoflurane-induced miR-211-5p promotes neuronal apoptosis by inhibiting Efemp2[J]. ASN Neuro, 2021, 13: 17590914211035036.
TIAN Y, CHEN K Y, LIU L D, et al. Sevoflurane exacerbates cognitive impairment induced by Aβ1-40 in rats through initiating neurotoxicity, neuroinflammation, and neuronal apoptosis in rat hippocampus[J]. Mediators Inflamm, 2018, 2018: 3802324.
ZHENG S Q, AN L X, CHENG X, et al. Sevoflurane causes neuronal apoptosis and adaptability changes of neonatal rats[J]. Acta Anaesthesiol Scand, 2013, 57(9): 1167-1174.
ZHANG L, XUE Z Y, LIU Q D, et al. Disrupted folate metabolism with anesthesia leads to myelination deficits mediated by epigenetic regulation of ERMN[J]. EBioMedicine, 2019, 43: 473-486.
SHI L L, MIAO Z J, CHENG Y Y, et al. Folic acid ameliorated sevoflurane exposure-induced decrease in differentiation capacity of oligodendrocyte precursor cells[J]. Anesthesiol Perioper Sci, 2024, 2(2): 13.
LIU B, XIN W, TAN J R, et al. Myelin sheath structure and regeneration in peripheral nerve injury repair[J]. Proc Natl Acad Sci U S A, 2019, 116(44): 22347-22352.
READHEAD C, POPKO B, TAKAHASHI N, et al. Expression of a myelin basic protein gene in transgenic shiverer mice: correction of the dysmyelinating phenotype[J]. Cell, 1987, 48(4): 703-712.
WU Z Y, XUE H, GAO Q S, et al. Effects of early postnatal sevoflurane exposure on oligodendrocyte maturation and myelination in cerebral white matter of the rat[J]. Biomed Pharmacother, 2020, 131: 110733.
ZHAO D, ZHANG M L, YANG L L, et al. GPR68 improves nerve damage and myelination in an immature rat model induced by sevoflurane anesthesia by activating cAMP/CREB to mediate BDNF[J]. ACS Chem Neurosci, 2022, 13(3): 423-431.
LIANG L R, ZENG T, ZHAO Y Y, et al. Melatonin pretreatment alleviates the long-term synaptic toxicity and dysmyelination induced by neonatal sevoflurane exposure via MT1 receptor-mediated Wnt signaling modulation[J]. J Pineal Res, 2021, 71(4): e12771.
ZUO Y, LI B W, XIE J H, et al. Sevoflurane anesthesia during pregnancy in mice induces cognitive impairment in the offspring by causing iron deficiency and inhibiting myelinogenesis[J]. Neurochem Int, 2020, 135: 104693.
ZHANG Z H, LIU H Q, JIA S S, et al. Spatial and temporal alterations of developing oligodendrocytes induced by repeated sevoflurane exposure in neonatal mice[J]. Biochem Biophys Res Commun, 2023, 640: 12-20.
SONG S Y, PENG K, MENG X W, et al. Single-nucleus atlas of sevoflurane-induced hippocampal cell type- and sex-specific effects during development in mice[J]. Anesthesiology, 2023, 138(5): 477-495.
ZHANG L, CHENG Y Y, XUE Z Y, et al. Sevoflurane impairs m6A-mediated mRNA translation and leads to fine motor and cognitive deficits[J]. Cell Biol Toxicol, 2022, 38(2): 347-369.
LI C, XIAO L, LIU X Y, et al. A functional role of NMDA receptor in regulating the differentiation of oligodendrocyte precursor cells and remyelination[J]. Glia, 2013, 61(5): 732-749.
MAURER U, CHARVET C, WAGMAN A S, et al. Glycogen synthase kinase-3 regulates mitochondrial outer membrane permeabilization and apoptosis by destabilization of MCL-1[J]. Mol Cell, 2006, 21(6): 749-760.
YANG F, ZHANG Y F, TANG Z Y, et al. Hemin treatment protects neonatal rats from sevoflurane-induced neurotoxicity via the phosphoinositide 3-kinase/Akt pathway[J]. Life Sci, 2020, 242: 117151.
GREEN D R, LLAMBI F. Cell death signaling[J]. Cold Spring Harb Perspect Biol, 2015, 7(12): a006080.
LIU X Y, JI J, ZHAO G Q. General anesthesia affecting on developing brain: evidence from animal to clinical research[J]. J Anesth, 2020, 34(5): 765-772.
SONG Q, MA Y L, SONG J Q, et al. Sevoflurane induces neurotoxicity in young mice through FAS/FASL signaling[J]. Genet Mol Res, 2015, 14(4): 18059-18068.
WERTZ I E, KUSAM S, LAM C, et al. Sensitivity to antitubulin chemotherapeutics is regulated by MCL1 and FBW7[J]. Nature, 2011, 471(7336): 110-114.
YON J H, DANIEL-JOHNSON J, CARTER L B, et al. Anesthesia induces neuronal cell death in the developing rat brain via the intrinsic and extrinsic apoptotic pathways[J]. Neuroscience, 2005, 135(3): 815-827.
CHEN X H, ZHOU X, LU D H, et al. Aberrantly expressed long noncoding RNAs are involved in sevoflurane-induced developing hippocampal neuronal apoptosis: a microarray related study[J]. Metab Brain Dis, 2016, 31(5): 1031-1040.
LI N N, ZHU R L, ZENG S, et al. The role of depolarizing activation of Na+-Ca2+ exchanger by oligodendrocyte progenitor cells in the effect of sevoflurane on myelination[J]. Life Sci, 2022, 308: 120951.
YOUNG J T, VLASOVA R M, HOWELL B R, et al. General anaesthesia during infancy reduces white matter micro-organisation in developing rhesus monkeys[J]. Br J Anaesth, 2021, 126(4): 845-853.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}