Mechanism of Fas-associated protein with death domain in promoting proliferation of head and neck squamous cell carcinoma cells
CHEN Yinan1, ZHENG Yang2, ZENG Hanlin1,3, LEI Ming,1
1.Shanghai Institute of Precision Medicine, Shanghai Ninth People's Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai 200125, China
2.Department of Oral Maxillofacial & Head and Neck Oncology, Shanghai Ninth People's Hospital, Shanghai Jiao Tong University School of Medicine; National Clinical Research Center for Oral Diseases; Shanghai Key Laboratory of Stomatology & Shanghai Research Institute of Stomatology, Shanghai 200011, China
3.Department of Oncology, Shanghai Ninth People's Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai 201999, China
目的·检测Fas相关死亡结构域蛋白(Fas-associated protein with death domain,FADD)在头颈部鳞状细胞癌(head and neck squamous cell carcinoma,HNSCC)中的表达水平,并探究FADD促进HNSCC细胞增殖的分子机制。方法·利用GEPIA 2数据库分析肿瘤组织中FADD表达水平及其与预后的关系;通过对HNSCC组织进行免疫组织化学染色(immunohistochemistry staining,IHC),探究FADD在正常、不典型增生和肿瘤组织中的表达水平变化;构建稳定低表达FADD的人HNSCC Fadu、HSC3细胞株,并通过蛋白印迹实验和实时荧光定量PCR(quantitative real-time PCR,qRT-PCR)方法进行验证;使用LiveCyte活细胞追踪系统、克隆形成、细胞活力检测等方法探究FADD对HNSCC细胞增殖水平的调控作用;使用免疫共沉淀串联质谱(co-immunoprecipitation mass spectrum,Co-IP/MS)鉴定与FADD发生相互作用的蛋白,并应用CRISPR/Cas9技术、LiveCyte活细胞追踪系统、蛋白印迹实验等方法对与FADD相互作用的蛋白进行进一步机制研究。结果·数据库分析显示FADD在头颈鳞癌中显著高表达,并与患者不良预后相关。免疫组化染色表明FADD在HNSCC患者正常组织、不典型增生及肿瘤组织中的表达水平呈现递增趋势。在HNSCC细胞中敲低FADD后,与对照组相比,细胞的增殖能力显著降低,形成克隆数减少。Co-IP/MS结果显示,FADD与CUT样同源盒1(CUT-like homeobox 1,CUX1)蛋白存在相互作用,敲低FADD后CUX1表达水平升高。同时,在HNSCC细胞中敲除CUX1能够显著促进肿瘤细胞增殖能力。敲除CUX1可部分逆转FADD低表达引起的增殖抑制。结论·FADD在HNSCC中具有显著促癌作用,并与不良预后相关。FADD可通过与CUX1发生相互作用降低其表达水平进一步调控肿瘤细胞的增殖能力。
关键词:Fas相关死亡结构域蛋白
;
CUT 样同源盒 1
;
头颈部鳞状细胞癌
;
细胞增殖
Abstract
Objective ·To detect the expression level of Fas-associated protein with death domain (FADD) in head and neck squamous cell carcinoma (HNSCC) and to explore the molecular mechanisms by which FADD promotes the proliferation of HNSCC cells. Methods ·The GEPIA 2 database was utilized to analyze the expression level of FADD in tumor tissues and to evaluate its association with prognosis. Immunohistochemistry staining (IHC) was performed on HNSCC tissues to investigate the changes in FADDexpression levels in normal, dysplastic, and tumor tissues. Stable FADD-knockdown Fadu and HSC3 cell lines were constructed and validated using Western blotting and quantitative real-time PCR (qRT-PCR). The regulatory effect of FADD on the proliferation of HNSCC cells was explored using the LiveCyte live-cell tracking system, colony formation assay, and cell viability assay. Proteins interacting with FADD were identified by co-immunoprecipitation mass spectrometry (Co-IP/MS), and further mechanistic studies were conducted using CRISPR/Cas9 technology, LiveCyte live-cell tracking system, and Western blotting. Results ·Analysis of the GEPIA2 database indicated that FADD was significantly overexpressed in head and neck cancer and was associated with poor prognosis. IHC staining showed that FADD expression levels progressively increased from normal to dysplastic to tumor tissues in HNSCC patients. Knockdown of FADD in HNSCC cells resulted in significantly reduced proliferation and colony formation compared to the control group. Co-IP/MS results showed that FADD interacted with the CUX1 protein, and FADD knockdown led to increased CUX1 expression. Moreover, CUX1 knockdown significantly promoted HNSCC cell proliferation and reversed the anti-proliferative phenotype caused by FADD knockdown. Conclusion ·FADD plays a significant pro-carcinogenic role in HNSCC and is associated with poor prognosis. FADD can further regulate tumor cell proliferation by interacting with CUX1 and suppressing its expression level.
Keywords:Fas-associated protein with death domain (FADD)
;
CUT-like homeobox 1 (CUX1)
;
head and neck squamous cell carcinoma (HNSCC)
;
cell proliferation
CHEN Yinan, ZHENG Yang, ZENG Hanlin, LEI Ming. Mechanism of Fas-associated protein with death domain in promoting proliferation of head and neck squamous cell carcinoma cells. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2025, 45(4): 404-414 doi:10.3969/j.issn.1674-8115.2025.04.002
头颈部肿瘤的发病率在全球常见恶性肿瘤中位居第六,2020年全球约有87万例新发病例,其中超过44万例死亡[1]。约有90%的头颈部肿瘤属于头颈部鳞状细胞癌(head and neck squamous cell carcinoma,HNSCC)。吸烟、饮酒及感染人乳头瘤病毒(human papillomavirus,HPV)是诱发HNSCC的高危因素[2-3]。目前主要采取手术、放疗及化疗的治疗方式,但由于HNSCC缺乏有效的早期诊断指标,复发转移率高且耐药,晚期HNSCC患者5年生存率仅为40%~50%[4-6]。因此,寻找HNSCC恶性进展的生物标志物,探索新的治疗靶点,对遏制肿瘤进展、提高HNSCC患者生存率具有重要意义。
Fas相关死亡结构域蛋白(Fas-associated protein with death domain,FADD)是死亡受体(death receptor,DR)介导的细胞凋亡途径中的重要受体蛋白。FADD蛋白由死亡结构域(death domain,DD)和死亡效应结构域(death effector domain,DED)组成。当FADD的DD与Fas或者肿瘤坏死因子受体1(tumor necrosis factor receptor 1,TNFR1)的DD相互作用后,能够通过N端的DED招募半胱天冬酶前体(pro-caspase),从而形成死亡诱导信号复合物(death-inducing signaling complex,DISC)。半胱天冬酶前体通过自身裂解的方式触发下游半胱天冬酶级联反应,促进细胞凋亡[7-8]。然而,近来有研究表明FADD还参与多种非细胞凋亡的过程,如调控细胞周期、细胞增殖、肿瘤侵袭及转移等[9-11]。FADD被证明是调控肿瘤增殖的重要因子,在肺癌、T淋巴母细胞淋巴瘤、胰腺癌的增殖中均发挥重要作用[12-14],但FADD调控肿瘤细胞增殖的具体机制需要进一步探索[15]。此外,FADD的表达还与肺腺癌、T淋巴母细胞淋巴瘤、阴茎鳞状细胞癌、非小细胞肺癌患者的不良预后相关[13,16-18],有望成为新型的预后标志物。
为检测FADD在HNSCC中的表达水平,本研究通过GEPIA2数据库分析肿瘤组织中FADD的表达水平,并利用HNSCC患者的病理切片进行免疫组织化学染色,检测FADD在正常组织、不典型增生及肿瘤组织中的表达水平。通过构建人HNSCC细胞系Fadu和HSC3 FADD敲低稳转株,在细胞水平上探究FADD对HNSCC细胞增殖能力的影响,并结合免疫共沉淀串联质谱(co-immunoprecipitation mass spectrum,Co-IP/MS)技术对FADD相互作用的调控蛋白进行分析,以阐明FADD促进HNSCC细胞增殖的分子机制。
1 材料与方法
1.1 实验材料
1.1.1 细胞系
人HNSCC Fadu细胞,人HNSCC HSC3细胞,人胚肾293T细胞,均购自American Type Culture Collection(ATCC)。使用添加10%胎牛血清的高糖DMEM培养液,于37 ℃、含5% CO2的细胞培养箱中培养。
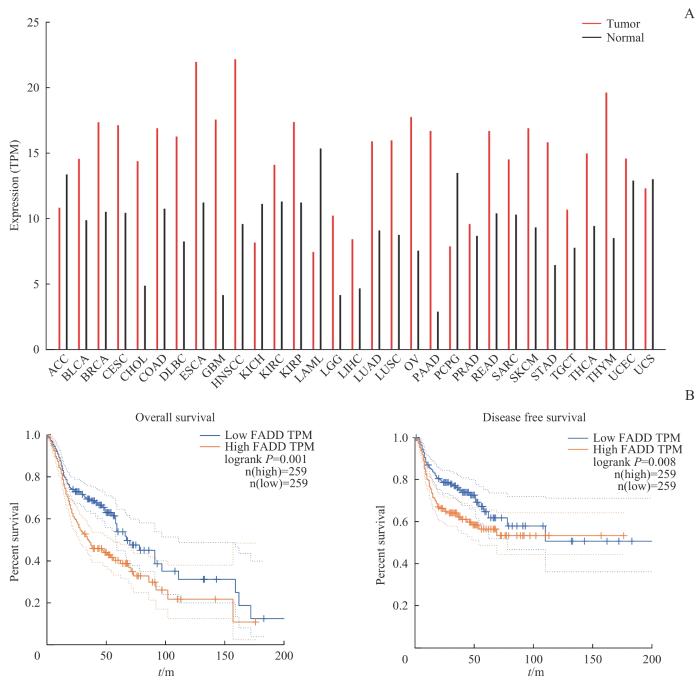
Note: A. FADD expression levels in different tumor types from the GEPIA 2 database. B. Relationship between FADD expression and prognosis in HNSCC patients based on the GEPIA 2 database. TPM—transcripts per million.
Fig 1
Expression levels of FADD in different cancers and its prognostic analysis
Note: A/B. HE (A) and IHC (B) staining of tumor sections from HNSCC patients. C. FADD H-scores in normal, dysplasia and tumor tissues. ①P<0.001, compared with normal tissues; ②P<0.001, compared with dysplastic tissues.
Fig 2
Relationship between FADD expression and HNSCC progression
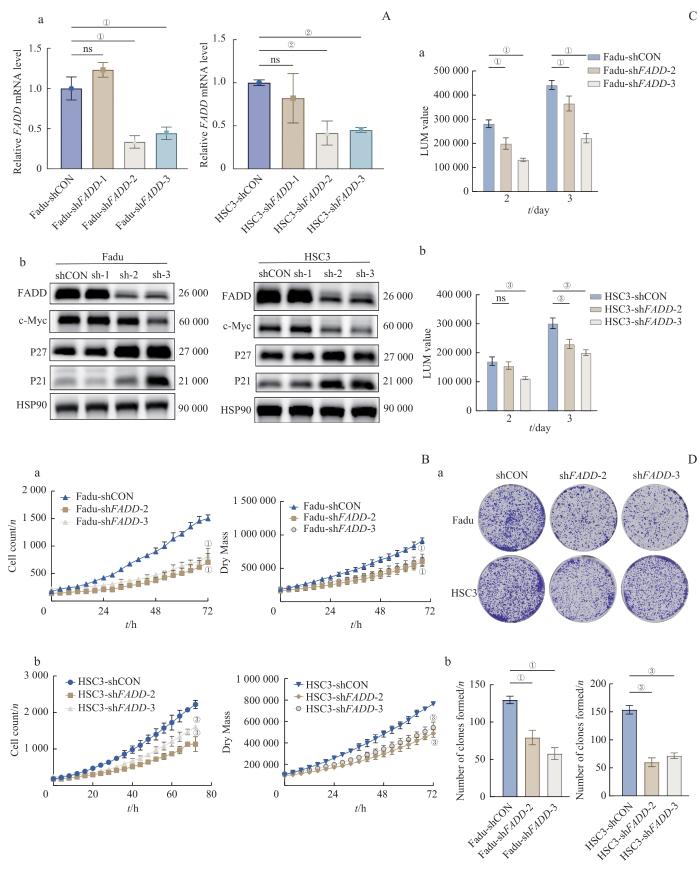
Note: A. Verification of FADD expression after stable knockdown of FADD in Fadu and HSC3 cells through qRT-PCR(a) and Western blotting (b). B. Cell count and dry mass curves for FadushFADD (a) and HSC3shFADD (b) detected by LiveCyte. C. Cell viability of FadushFADD (a) and HSC3shFADD (b). D. Plate colony formation assay in Fadu and HSC3 cells after FADD knockdown. ①P<0.001, ②P=0.005, ③P<0.001.
Fig 3
FADD knockdown inhibited the proliferation of HNSCC cells
Note: A. Workflow of Co-IP/MS samples preparation. B. Samples reserved for Western blotting. C/D. Heatmap of potential FADD-interacting proteins in Fadu (C) and HSC3 cell lines (D). Red, white and blue respectively represent high, medium and low levels of relative protein expression. CUX1 is highlighted in red in both Fadu and HSC3 cell lines. E/F. The Western blotting results of CUX1 protein detection in Co-IP samples (E) and FADD-knockdown HNSCC cells (F).
Note: A. Detection of CUX1 expression in HSC3 and Fadu cells after CUX1 knockout by Western blotting. B. Plate colony formation assay in FadusgCUX1 and HSC3sgCUX1 cells. C. Cell viability of FadusgCUX1 and HSC3sgCUX1 cells. D. Detection of P21 after CUX1 knockout in HSC3 and Fadu cells by Western blotting. E. The effect of CUX1 knockout on proliferation in stable-FADD-knockdown Fadu and HSC3 cells. ①P=0.004, ②P=0.008, ③P=0.001, ④P=0.001; ⑤P<0.001, compared with the FaduMOCK group; ⑥P<0.001 compared with the HSC3MOCK group.
Fig 5
FADD regulates the proliferation of HNSCC cells through CUX1
CHEN Yinan, ZHENG Yang, ZENG Hanlin and LEI Ming designed the whole research project. CHEN Yinan mainly completed the work of experiment execution, data analysis and writing of the manuscript. ZHENG Yang and ZENG Hanlin were mainly responsible for the guidance of experimental scheme and data analysis, as well as the revision of the manuscript. All authors have read the final manuscript and approved the submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interest.
SUNG H, FERLAY J, SIEGEL R L, et al. Global cancer statistics 2020: globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries[J]. CA Cancer J Clin, 2021, 71(3): 209-249.
SUN Z, SUN X D, CHEN Z W, et al. Head and neck squamous cell carcinoma: risk factors, molecular alterations, immunology and peptide vaccines[J]. Int J Pept Res Ther, 2022, 28(1): 19.
WONG K C W, JOHNSON D, HUI E P, et al. Opportunities and challenges in combining immunotherapy and radiotherapy in head and neck cancers[J]. Cancer Treat Rev, 2022, 105: 102361.
AWADIA S, SITTO M, RAM S, et al. The adapter protein FADD provides an alternate pathway for entry into the cell cycle by regulating APC/C-Cdh1 E3 ubiquitin ligase activity[J]. J Biol Chem, 2023, 299(6): 104786.
PATTJE W J, MELCHERS L J, SLAGTER-MENKEMA L, et al. FADD expression is associated with regional and distant metastasis in squamous cell carcinoma of the head and neck[J]. Histopathology, 2013, 63(2): 263-270.
WEI S Y, CHEN Z G, LING X Y, et al. Comprehensive analysis illustrating the role of PANoptosis-related genes in lung cancer based on bioinformatic algorithms and experiments[J]. Front Pharmacol, 2023, 14: 1115221.
MARÍN-RUBIO J L, PÉREZ-GÓMEZ E, FERNÁNDEZ-PIQUERAS J, et al. S194-P-FADD as a marker of aggressiveness and poor prognosis in human T-cell lymphoblastic lymphoma[J]. Carcinogenesis, 2019, 40(10): 1260-1268.
ZHANG R, LIU Y T, HAMMACHE K, et al. The role of FADD in pancreatic cancer cell proliferation and drug resistance[J]. Oncol Lett, 2017, 13(3): 1899-1904.
ZHENG Y, SHENG S, MA Y, et al. FADD amplification is associated with CD8+ T-cell exclusion and malignant progression in HNSCC[J]. Oral Dis, 2024, 30(8): 5007-5021.
HE M, HE Y Y, XU J, et al. Upregulated FADD is associated with poor prognosis, immune exhaustion, tumor malignancy, and immunotherapy resistance in patients with lung adenocarcinoma[J]. Front Oncol, 2023, 13: 1228889.
XUE T, YAN R, LI Z S, et al. Prognostic significance and immune correlates of FADD in penile squamous cell carcinoma[J]. Virchows Arch, 2023, 482(5): 869-878.
CIMINO Y, COSTES A, DAMOTTE D, et al. FADD protein release mirrors the development and aggressiveness of human non-small cell lung cancer[J]. Br J Cancer, 2012, 106(12): 1989-1996.
LIVINGSTON S, CARLTON C, SHARMA M, et al. Cux1 regulation of the cyclin kinase inhibitor p27kip1 in polycystic kidney disease is attenuated by HDAC inhibitors[J]. Gene X, 2019, 2: 100007.
ITOH N, YONEHARA S, ISHII A, et al. The polypeptide encoded by the cDNA for human cell surface antigen Fas can mediate apoptosis[J]. Cell, 1991, 66(2): 233-243.
LIU Y B, FAN C X, ZHANG Y F, et al. RIP1 kinase activity-dependent roles in embryonic development of Fadd-deficient mice[J]. Cell Death Differ, 2017, 24(8): 1459-1469.
CHEN G A, BHOJANI M S, HEAFORD A C, et al. Phosphorylated FADD induces NF-kappaB, perturbs cell cycle, and is associated with poor outcome in lung adenocarcinomas[J]. Proc Natl Acad Sci USA, 2005, 102(35): 12507-12512.
GONZÁLEZ-MOLES M Á, AYÉN Á, GONZÁLEZ-RUIZ I, et al. Prognostic and clinicopathological significance of FADD upregulation in head and neck squamous cell carcinoma: a systematic review and meta-analysis[J]. Cancers, 2020, 12(9): 2393.
EYTAN D F, SNOW G E, CARLSON S, et al. SMAC mimetic birinapant plus radiation eradicates human head and neck cancers with genomic amplifications of cell death genes FADD and BIRC2[J]. Cancer Res, 2016, 76(18): 5442-5454.
CHIEN H T, CHENG S D, CHUANG W Y, et al. Clinical Implications of FADD Gene Amplification and Protein Overexpression in Taiwanese Oral Cavity Squamous Cell Carcinomas[J]. PLOS One, 2016, 11(10): e0164870.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}