

尽管5-HT的重要性已被广泛认可,其在中枢及外周的具体作用机制仍远未完全阐明。这一现状部分归因于5-HT检测技术的局限性。目前常用的5-HT检测方法包括微量渗析法[11]、电生理技术[12]和快速扫描循环伏安法[13]等。然而这些方法存在时空分辨率低、特异性差、灵敏度不足及侵入性强等局限性[14],无法实现对5-HT动态释放的实时、灵敏追踪,因而限制了5-HT递质释放机制的研究。近年来,基因编码的5-HT荧光探针因其能高时空分辨率地检测活体动物脑内5-HT的动态变化,为体内外5-HT检测提供了新的手段。以细菌周质结合蛋白(periplasmic binding proteins,PBPs)为骨架开发的iSeroSnFR1.0[15],是第一个以PBPs为骨架开发的基因编码5-HT荧光探针。当5-HT与PBPs结合时,会引起后者构象变化,并将该变化传导至荧光报告位点构象敏感的循环重排荧光蛋白(circular permutated fluorescence proteins,cpFP),影响其发色团周围的微环境,从而引起荧光强度变化。
然而,我们前期研究发现,iSeroSnFR1.0的基底荧光强度较弱,在培养的海马脑片中,表达该探针的细胞膜成像效果不理想,导致边界不清晰,限制了其对内源性5-HT信号的追踪能力[16]。同样,基于PBPs开发并进一步优化后的谷氨酸递质探针SF-iGluSnFR,在培养神经元中的衰减动力学快于
钙离子探针GCaMP6f[17],这是iSeroSnFR1.0目前还无法实现的。因此,本研究拟进一步优化iSeroSnFR1.0以提升其基底荧光强度、反应动态范围及动力学性能,使其能够更适用于快速检测内源性5-HT信号。这一优化不仅可为科研人员在不同研究需求下选择合适的5-HT荧光探针提供更丰富的工具选择,同时也为解析神经系统及其他系统疾病中5-HT异常释放的机制提供新的研究思路。
1 材料与方法
1.1 材料
1.1.1 实验动物
本实验选用胚胎发育第17日(E17)或出生第1日(P0)的C57BL/6小鼠,用于原代皮层神经元的提取和培养。小鼠购自上海灵畅生物科技有限公司,在上海交通大学医学院动物科学实验中心(SPF级)饲养。实验动物生产许可证号SCXK(沪)2023-0010,实验动物使用许可证号SYXK(沪)2023-0041。饲养环境温度为24 ℃±1 ℃,湿度控制在55%,并实行12 h光暗循环,允许小鼠自由进食和饮水。
1.1.2 荧光探针
iSeroSnFR1.0、iAChSnFR荧光探针及Sindbis病毒载体由美国弗吉尼亚大学J. Julius ZHU教授实验室提供,病毒包装在上海交通大学医学院完成。
1.1.3 主要试剂及仪器
Gibson Assembly Master Mix(New England Biolabs,美国),Q5 High-Fidelity DNA Polymerase(New England Biolabs,美国),mMESSAGE mMACHINE(Invitrogen,美国),5-羟色胺盐酸盐(Sigma-Aldrich,美国),DMEM培养基(Gibco,美国),Lipo8000(碧云天),Neurobasal-A培养基(Gibco,美国),B27(Gibco,美国)。正置显微镜(Leica,德国),PCR仪(Thermo Fisher Scientific,美国),放大器MultiClamp 700B(Molecular devices,美国),生物刺激隔离器(AMPI,以色列)。
1.2 实验方法
1.2.1 质粒构建
首先,分别设计靶向iAChSnFR的cpsfGFP基因和iSeroSnFR1.0骨架PBPs基因的特异性引物,利用Q5 High-Fidelity DNA Polymerase进行PCR扩增,以获得带有同源末端的目标片段。接着,使用Gibson Assembly Master Mix对这2个片段进行基因重组,使得iSeroSnFR1.0中的原cpsfGFP基因被iAChSnFR中的cpsfGFP基因所替换,从而获得新的基因编码的5-HT荧光探针iSeroSnFR1.2。
1.2.2 Sindbis病毒包装
携带iSeroSnFR1.0和iSeroSnFR1.2基因序列的质粒经过XbaⅠ和PmlⅠ限制性内切酶双酶切后,将目的片段克隆至Sindbis病毒载体pSinRep5中。随后,使用PacⅠ内切酶线性化质粒,并利用体外转录试剂盒mMESSAGE mMACHINE在37 ℃条件下进行体外转录,以合成病毒RNA。将合成的RNA通过电穿孔法转染BHK-21细胞,转染约48 h后收集含病毒颗粒的细胞培养上清液,高速离心后浓缩1 000倍并保存[18-19]。
1.2.3 HEK293细胞培养、质粒转染和加药反应
将本研究使用的HEK293细胞加入含10%胎牛血清和1%青链霉素的DMEM培养基中,置于37 ℃、5% CO₂的恒温培养箱中进行培养。采用Lipo8000脂质体将携带探针序列的质粒转染入HEK293细胞中。转染后培养20~24 h,通过滴加5-HT进行成像,并观察荧光响应,使用正置荧光显微镜进行检测。
1.2.4 原代小鼠皮层神经元提取、培养及病毒感染
本研究中使用的神经元为E17或P0时期的C57BL/6小鼠胚胎大脑皮层神经元。首先,处死孕鼠后取出胚胎(对于P0小鼠,则无需此步骤),在无菌条件下分离皮层组织。使用0.25%胰蛋白酶在37 ℃下消化15 min,然后将组织分散为单个细胞。细胞悬液在预冷的含有10%胎牛血清和2 mmol/L谷氨酰胺的Neurobasal培养基中以1×106个/mL的密度接种到预先用L-赖氨酸包被的24孔板中,培养4 h后更换为无血清培养基。在37 ℃、5% CO₂的条件下培养,每5 d更换1次培养基。在体外培养第13日时加入携带探针的Sindbis病毒,表达18 h后进行电刺激实验。所有实验操作均在无菌条件下进行,以确保细胞培养的质量和一致性。
1.2.5 HEK293细胞瞬时给药(puff)和荧光成像同步实验
将成功表达探针的HEK293细胞培养玻片转移到记录槽中。使用含有5 mmol/L 5-HT的玻璃微电极,靠近单个细胞的膜表面进行瞬时给药,持续给药时间为10 ms,帧间隔时间为10 ms,记录总时长为10 s。同时,使用Hamamatsu ORCA-Flash 4.0相机进行荧光成像,激发光波长λ=460 nm。为了确保图像捕获与药物喷洒同步,相机设置为外部触发模式。
1.2.6 原代小鼠皮层神经元电刺激诱导5-HT释放和荧光成像同步实验
将成功表达探针的皮层神经元培养玻片转移到记录槽中。随后,将双极刺激电极接触到阳性细胞附近的神经元上,刺激电压设置为3~5 mV,持续刺激时间为0.4 ms,帧间隔时间为10 ms,记录总时长为10 s。同时,使用Hamamatsu ORCA-Flash 4.0相机进行荧光成像,激发光波长λ=460 nm。为了确保图像捕获与电刺激同步,相机设置为外部触发模式。
1.3 统计学分析
使用HCImage Live和Matlab软件进行图像处理,并利用Igor及GraphPad Prism 9.0软件进行数据分析。2组数据之间的比较采用双尾独立样本t检验。所有统计数据均以x±s表示。P<0.05表示差异具有统计学意义。
2 结果
2.1 iSeroSnFR1.0的优化及性能表征
研究显示,荧光蛋白的氨基酸序列变化会显著影响其折叠过程,从而改变荧光亮度和光稳定性等性能[20]。基于这一原理,为提高iSeroSnFR1.0探针的性能,我们采用Gibson组装法,将基底荧光强度较高的乙酰胆碱探针iAChSnFR中的cpsfGFP基因序列替换到iSeroSnFR1.0中,从而构建出新的5-HT探针iSeroSnFR1.2。氨基酸序列分析显示,iSeroSnFR1.2的cpsfGFP中有3个突变位点,其中2个位于PBPs与cpsfGFP相互作用界面附近(图1A)。这提示这些突变可能对探针的构象变化起重要作用。
图1

图1
5-HT荧光探针iSeroSnFR1.2在HEK293细胞上的优化与表征
Note: A. Schematic illustration of the optimization strategy for iSeroSnFR1.0. The cpsfGFP sequence from iAChSnFR was transplanted into iSeroSnFR1.0 to generate iSeroSnFR1.2, with key amino acid changes highlighted. B. Fluorescence response changes. Scale bar: 50 μm. C. Normalized values of iSeroSnFR1.0- and iSeroSnFR1.2-expressing HEK293 cells treated with 250 μmol/L 5-HT, ①P=0.012 (n=10 cells from 3 independent experiments). D. Normalized basal fluorescence values of HEK293 cells expressing iSeroSnFR1.0 and iSeroSnFR1.2, ②P<0.001 (n=26 cells from 3 independent experiments). E. Normalized ΔF/F0 values of the iSeroSnFR1.2 in response to various neurotransmitters (250 μmol/L 5-HT, 20 μmol/L ACh, 20 μmol/L DA, 20 μmol/L Epi, 20 μmol/L GABA, 20 μmol/L Glu, 20 μmol/L His), ③P<0.001 (n=10 cells from 3 independent experiments). Statistical analysis was performed using a two-tailed unpaired
Fig 1
Optimization and characterization of iSeroSnFR1.2 in HEK293 cells
为了评估iSeroSnFR1.2对外源性5-HT的检测能力,我们在HEK293细胞中过表达iSeroSnFR1.0和iSeroSnFR1.2,并给予5 mmol/L 5-HT药物处理并观察反应。结果显示,iSeroSnFR1.2对5-HT的荧光响应显著高于iSeroSnFR1.0(图1B、C),同时iSeroSnFR1.2不仅提高了探针的荧光响应,也增加了细胞的基底荧光强度(图1D)。这一特性有助于在高背景环境中进行荧光成像。为了进一步评估iSeroSnFR1.2对5-HT的特异性,我们在表达iSeroSnFR1.2的HEK293细胞中进行了其他神经递质的药物处理实验。结果表明,乙酰胆碱(acetylcholine,ACh)、多巴胺(dopamine,DA)、肾上腺素(epinephrine,Epi)、γ-氨基丁酸(γ-aminobutyric acid,GABA)、谷氨酸(glutamate,Glu)及组胺(histamine,His)等神经递质均未能诱导可检测的荧光反应(图1E)。这些初步结果表明,经过优化的iSeroSnFR1.2相较于iSeroSnFR1.0在检测外源性5-HT时表现出更强的基底荧光和荧光响应,并且对5-HT仍保留较高的分子特异性。
2.2 iSeroSnFR1.2检测外源性5-HT的荧光反应和动力学
为了评估iSeroSnFR1.2对外源性5-HT的荧光响应幅度和动力学特征,我们在培养的HEK293细胞中过表达探针,随后使用含有5 mmol/L 5-HT的玻璃微电极靠近单个细胞的膜表面进行瞬时给药实验(图2A)。我们记录了其荧光反应并绘制出荧光反应曲线(图2C)。通过曲线拟合计算上升时间(从最大响应的10%上升到90%的时间)和衰减时间常数(从最大响应下降到最大响应的50%的时间),以表征其动力学特性,并对iSeroSnFR1.2与iSeroSnFR1.0的性能进行比较分析。实验结果表明,表达iSeroSnFR1.2的细胞不仅表现出更强的基底荧光,其对5-HT的荧光响应也显著增强(图2B)。具体而言,iSeroSnFR1.2的最大荧光响应为iSeroSnFR1.0的1.5倍(图2D);在动力学方面,iSeroSnFR1.2对5-HT反应的上升时间为36.3 ms,略快于iSeroSnFR1.0(为44.9 ms)(图2E)。更显著的是,iSeroSnFR1.2的荧光衰减时间常数为1 003.6 ms,较iSeroSnFR1.0(1 730.4 ms)减少了42.0%(图2F)。综上所述,在HEK293细胞中,iSeroSnFR1.2相比于iSeroSnFR1.0显著提高了对外源性5-HT的响应幅度,并表现出更快的动力学特性,特别是在信号衰减方面。这些改进使得iSeroSnFR1.2成为一个更灵敏、高时间分辨率的5-HT荧光探针,具有快速检测5-HT动态改变的潜力。
图2
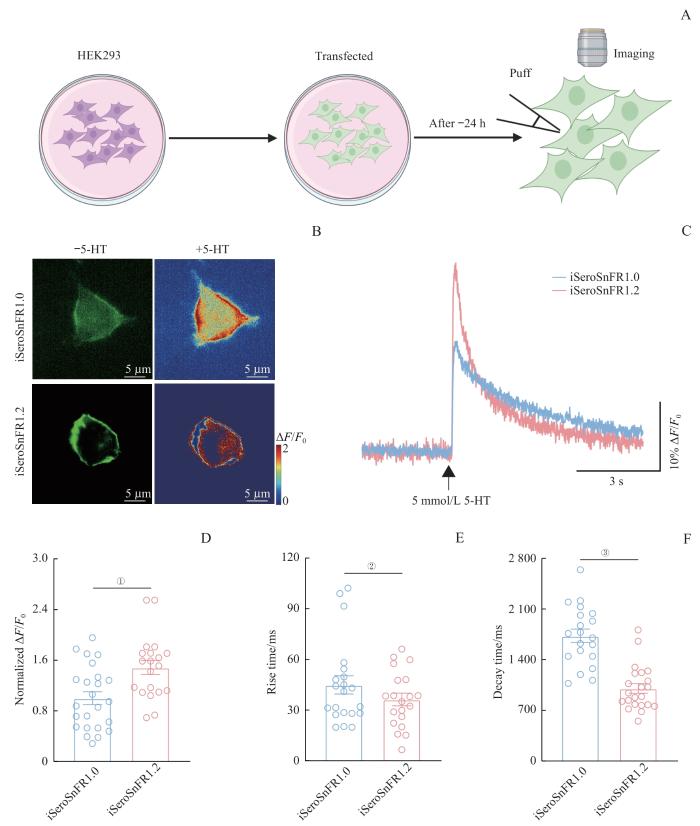
图2
5-HT荧光探针iSeroSnFR1.2在HEK293细胞中检测外源性5-HT的表征
Note: A. Schematic drawing outlining the design of puff experiments in HEK293 cells. B. Representative fluorescence images of HEK293 cells expressing iSeroSnFR1.0 or iSeroSnFR1.2. Left: basal fluorescence; Right: heat maps showing fluorescence response to 5 mmol/L 5-HT perfusion. Scale bar: 5 μm. C. Normalized fluorescence response of HEK293 cells expressing iSeroSnFR1.0 or iSeroSnFR1.2 to a brief puff (10 ms) of 5 mmol/L 5-HT. D. Normalized values comparing fluorescence responses between iSeroSnFR1.0 and iSeroSnFR1.2. ①P=0.003 (n=20 cells from 3 independent experiments). E. Comparison of rise times of responses to the 5 mmol/L 5-HT puff between iSeroSnFR1.0 and iSeroSnFR1.2. ②P=0.461 (n=20 cells from 3 independent experiments). F. Comparison of decay times between iSeroSnFR1.0 and iSeroSnFR1.2. ③P<0.001 (n=22 cells from 3 independent experiments). Statistical analysis was performed using a Two-tailed unpaired Student′s t-test.
Fig 2
Characterization of iSeroSnFR1.2′s response to exogenous 5-HT in HEK293 cells
2.3 iSeroSnFR1.2检测内源性5-HT释放的荧光响应和动力学
为了进一步探究iSeroSnFR1.2对内源性5-HT的荧光响应幅度和动力学特征,并验证其在不同实验条件下的适用性,我们采用携带iSeroSnFR1.0和iSeroSnFR1.2的Sindbis病毒感染小鼠皮层神经元。18 h后,在表达探针的阳性细胞附近施加1个电脉冲刺激(持续时间0.4 ms),以诱导神经元释放内源性5-HT(图3A),并记录其荧光反应并绘制出荧光反应曲线(图3C)。结果显示,2种探针均能检测电刺激诱导的神经元内源性5-HT的释放(图3B)。具体而言,在荧光响应大小方面,iSeroSnFR1.2的荧光反应为iSeroSnFR1.0的2.7倍(图3D);在动力学特性方面,iSeroSnFR1.2表现出更快的响应速度,其上升时间为429.4 ms,比iSeroSnFR1.0(766.2 ms)缩短44.0%(图3E);衰减时间常数为814.6 ms,比iSeroSnFR1.0 (1 111.2 ms)缩短26.7%(图3F)。
图3
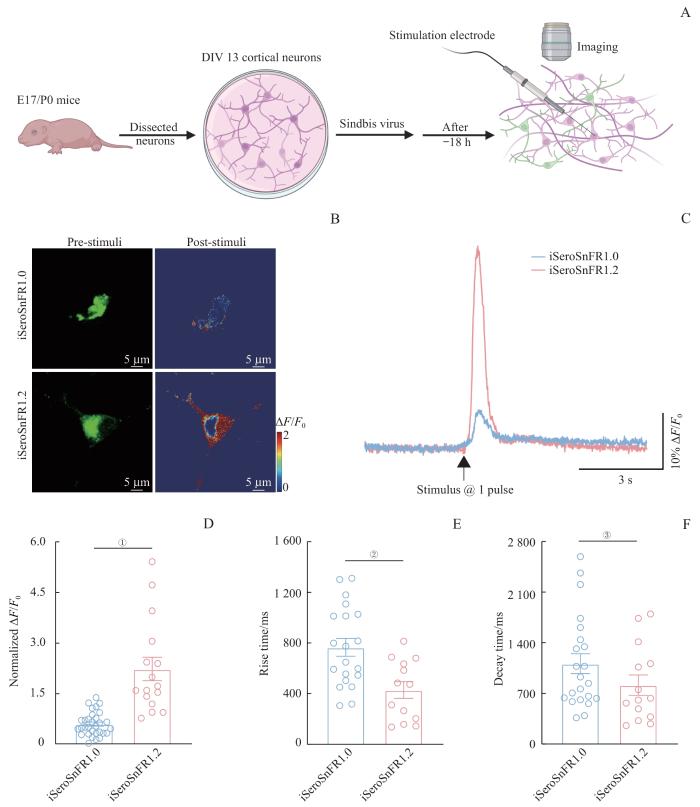
图3
5-HT荧光探针iSeroSnFR1.2检测培养皮层神经元中内源性5-HT的释放
Note: A. Schematic illustration outlines the design of electrical stimulation experiments in mouse cortical neurons. B. Representative fluorescence imaging of cortical neurons expressing either iSeroSnFR1.0 or iSeroSnFR1.2. Images display GFP pseudocolor and corresponding heatmaps before (left) and after (right) a single 2-second electrical stimulus. Scale bar: 5 μm. The stimulus consisted of a single pulse. C/D. Simultaneous fluorescence responses (C) and normalized values (D) of iSeroSnFR1.0- and iSeroSnFR1.2-expressing mouse cortical neurons in response to electrical stimulation at a single pulse. ①P<0.001 (n=16 from 3 independent experiments). E. Comparison of rise times in responses to electrical stimulation between iSeroSnFR1.0 and iSeroSnFR1.2. ②P=0.003 (n=13 from 3 independent experiments). F. Comparison of decay times in response to electrical stimulation between iSeroSnFR1.0 and iSeroSnFR1.2. ③P=0.032 (n=14 from 3 independent experiments). Statistical analysis was performed using a two-tailed unpaired
Fig 3
Detection of endogenous 5-HT release in cultured cortical neurons using iSeroSnFR1.2
这些在培养神经元中的数据进一步证实,iSeroSnFR1.2能够有效检测电刺激诱导的内源性5-HT释放。此外,与iSeroSnFR1.0相比,iSeroSnFR1.2在检测内源性5-HT时展现出更显著的响应动态范围和更快的动力学特性。这些结果表明,iSeroSnFR1.2是一个更为灵敏和具有高时间分辨率的5-HT荧光探针。期待在各种实验条件下均能提供更准确的5-HT动态信息。
3 讨论
本研究利用DNA重组技术将iAChSnFR的cpsfGFP基因序列整合到iSeroSnFR1.0中,成功构建了新型5-HT荧光探针iSeroSnFR1.2。随后在HEK293细胞和培养的小鼠皮层神经元中对该探针进行了评估,结果表明:iSeroSnFR1.2在检测5-HT方面显著优于iSeroSnFR1.0,尤其在荧光反应大小和动力学特性方面表现突出。值得注意的是,iSeroSnFR1.2不仅提高了荧光反应幅度,还增强了基底荧光强度,这一特性显著提升了探针在高背景环境中的成像稳定性和可靠性。在优化过程中,我们注意到乙酰胆碱探针iAChSnFR的基底荧光强度显著高于iSeroSnFR1.0。考虑到基底荧光强度与荧光蛋白的内在特性密切相关[21],我们将iSeroSnFR1.0中的荧光蛋白cpsfGFP替换为iAChSnFR中相应的荧光蛋白基因。实验结果证实,这一改造使新型探针iSeroSnFR1.2的基底荧光强度显著提升,在培养神经元中是iSeroSnFR1.0的1.2倍,预计可能大幅提高该探针在培养脑片和体内实验中的成像性能。更深入的分析揭示,替换后的cpsfGFP序列中存在I107T和Y123H 2个关键突变。这2个位点位于荧光蛋白与PBPs的相互作用区域,可能显著影响蛋白质-配体相互作用的构象变化过程从而改变cpsfGFP蛋白折叠发光[22]。我们的实验数据验证了上述假设,观察到荧光信号的上升时间和衰减时间常数明显缩短,同时基底荧光强度和荧光反应显著增大。这些变化可能暗示配体结合口袋的动态特性发生了实质性改变,从而优化了探针的整体性能。
在本研究中,我们发现iSeroSnFR1.2在HEK293细胞中的上升时间与iSeroSnFR1.0之间无显著差异(图2E),但在培养皮层神经元中表现出显著差异(图3E)。这可能是由于HEK293细胞表面具有黏性物质,并且给药过程速度较快,导致在puff给药时膜表面的探针与5-HT接触不充分,而电刺激神经元诱导内源性5-HT释放则几乎不存在该问题。此外,iSeroSnFR1.2对电刺激诱导神经元内源性5-HT的上升时间平均值为429.4 ms。基于PBPs开发的第一个神经递质探针——谷氨酸探针iGluSnFR在经过多轮优化后得到的iGluSnFR3,其对电刺激诱导神经元内源性谷氨酸的上升时间为20.0 ms[23],已经接近可以识别单个动作电位的时间分辨率。同时,基于PBPs开发的乙酰胆碱探针iAChSnFR在急性脑片中的纹状体中检测乙酰胆碱的上升时间为37.2 ms[24]。而iSeroSnFR1.2响应5-HT的上升时间是以上2种基因编码的神经递质荧光探针的数十倍,这表明其在动力学上还有待进一步提高。
2024年,北京大学李毓龙实验室[25]基于GRAB5-HT1.0进行多位点突变和筛选,成功优化开发得到GRAB5-HT3.0。从动力学特征、亲和力及信噪比等多个方面进行了表征。结果显示,在HEK293细胞和培养神经元中GRAB5-HT3.0的亲和力分别是GRAB5-HT1.0的3.0倍和7.5倍,在培养神经元中GRAB5-HT3.0的荧光响应是GRAB5-HT1.0的6.8倍。此外,进一步在活体动物中验证发现,GRAB5-HT3.0能够检测行为小鼠在睡眠觉醒、癫痫发作等行为过程的体内5-HT动态变化过程。尽管我们已经在HEK293以及培养神经元中证明iSeroSnFR1.2相比于iSeroSnFR1.0在多个方面显著改善,但是相比于GRAB5-HT3.0在各方面提升的幅度,iSeroSnFR1.2仍然具有较大的提升空间。此外,iSeroSnFR1.2缺乏在脑片及行为小鼠中的进一步验证。
我们观察到,这几类探针在优化过程中均采取易错性PCR、重组或定点随机突变的方式扩大突变体文库,并筛选到最终性能表现优良的神经递质探针。鉴于此发现,以及PBPs的可溶性特性及其在结构上的显著适应性[26-27],下一步计划将利用蛋白质工程策略,例如定向进化技术,对iSeroSnFR1.2的PBPs结构部分进行理性设计,以进一步实现对5-HT亲和力和结合速率与解离动力学的优化。这种策略有望产生具有更高灵敏度、更快动力学特性或更广动态范围的新一代5-HT探针。此外,在探针应用方面,近年来基于基因编码探针与图像分析算法融合的技术突破,为5-HT研究提供了新维度——通过构建基因编码探针和图像处理算法系统,研究者不仅实现了5-HT动态分布的可视化追踪,而且精准解析了递质释放模式;该方法具有广阔的应用前景[28]。
综上所述,iSeroSnFR1.2在检测5-HT的基底荧光强度、荧光反应大小和动力学特性方面均显著优于iSeroSnFR1.0。这些优化使其成为一种有潜力的工具。未来的研究将继续优化探针性能,进一步探索iSeroSnFR1.2与多种分析手段结合应用于5-HT相关研究,为深入解析神经系统及其他生理系统中的5-HT动态调控机制提供新的研究工具。未来,该探针有望在神经科学、药理学及相关领域发挥重要作用。
作者贡献声明
徐慕凡、张坤、王静怡和高欣柯参与实验实施和数据分析;程傲冰、张鹏、徐慕凡、张坤参与论文写作和修改;程傲冰、张鹏负责课题设计与实验指导;所有作者均阅读并同意最终稿件的提交。
AUTHOR's CONTRIBUTIONS
XU Mufan, ZHANG Kun, WANG Jingyi and GAO Xinke performed the experiments and analyzed the data. CHENG Aobing, ZHANG Peng, XU Mufan, and ZHANG Kun wrote and revised the manuscript. CHENG Aobing and ZHANG Peng conceived, designed and supervised the project. All authors have read the last version of paper and consented to its submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
参考文献

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}