Study on the mechanism of KRAS R68G secondary mutation-induced resistance to KRASG12D-targeted inhibitor MRTX1133
WANG Gaoming1,2,3, CUI Ran4, LI Yanjing,2,3, LIU Yingbin,1,2,3
1.Department of Biliary-Pancreatic Surgery, Renji Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai 200127, China
2.State Key Laboratory of Systems Medicine for Cancer, Shanghai Cancer Institute, Renji Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai 200127, China
3.Shanghai Key Laboratory for Cancer Systems Regulation and Clinical Translation (CSRCT-SHANGHAI), Renji Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai 200127, China
4.Department of Hepatopancreatobiliary Surgery, East Hospital Affiliated Tongji University, Shanghai 200120, China
目的·从原子层面探索KRASG12D/R68G突变诱发肿瘤细胞对MRTX1133耐药的机制。方法·从RCSB蛋白质数据库(Protein Data Bank,PDB)获取KRASG12D与MRTX1133相互作用复合物的晶体结构数据。使用PyMOL软件将KRAS第68位的精氨酸突变为甘氨酸(R68G),构建KRASG12D和KRASG12D/R68G分别与MRTX1133相互作用体系的初始构象。使用LEaP程序构建带有周期性边界的模拟体系,应用ff19SB力场计算KRAS中标准氨基酸的力场参数,应用GAFF(general AMBER force field)力场计算MRTX1133的力场参数,应用TIP3P(intermolecular potential three point)力场计算水分子的力场参数。使用Amber软件对体系进行能量最小化,体系升温至300 K后,进行等温等容平衡和等温等压运动的计算。使用cpptraj轨迹分析软件计算每个体系的均方根偏差(root mean square deviation,RMSD)、体系中每个氨基酸的均方根波动(root mean square fluctuation,RMSF),对轨迹进行主成分分析(principal component analysis,PCA),计算MRTX1133和GDP的溶剂可及表面积(solvent-accessible surface area,SASA)。测量区域之间氢键形成的数量,并计算氨基酸之间的动态交叉相关矩阵(dynamic cross-correlation matrix,DCCM)。结果·RMSD分析显示KRASG12D/R68G体系中KRAS的变化幅度大于KRASG12D体系。RMSF分析显示KRASG12D/R68G体系中KRAS的Switch Ⅰ和Switch Ⅱ区域的波动幅度明显大于KRASG12D体系。PCA分析提示KRASG12D/R68G体系中KRAS的Switch Ⅰ和Switch Ⅱ区域更多地处于向外打开的状态。两体系中Switch Ⅰ与P-loop之间距离以及Switch Ⅱ与P-loop之间距离的比较显示了KRASG12D/R68G体系中的GDP和MRTX1133的结合口袋与KRASG12D体系相比均显著扩大。SASA分析显示KRASG12D/R68G体系中的GDP和MRTX1133的溶剂暴露面积与KRASG12D体系相比均明显增加。DCCM分析显示KRASG12D/R68G体系中Switch Ⅰ、Switch Ⅱ和P-loop区域之间存在更多的分离运动。结论·KRASG12D/R68G突变破坏了Switch Ⅰ和Switch Ⅱ区域之间的相互作用,导致了Switch Ⅰ和Switch Ⅱ的分离,继而导致MRTX1133的结合口袋开放,增加了MRTX1133的溶剂暴露面积,从而加速了MRTX1133的解离,最终导致KRASG12D/R68G对MRTX1133耐药。
关键词:MRTX1133
;
KRAS继发突变
;
分子动力学模拟
;
耐药机制
Abstract
Objective ·To explore the mechanism at the atomic level by which the KRASG12D/R68G mutation induces tumor cell resistance to MRTX1133. Methods ·The crystal structure data of the KRASG12D-MRTX1133 complex were obtained from the RCSB Protein Data Bank (PDB). PyMOL software was used to mutate arginine at position 68 of KRAS to glycine (R68G), constructing the initial conformations of the KRASG12D-MRTX1133 and KRASG12D/R68G-MRTX1133 complexes. The LEaP module was used to build simulation systems under periodic boundary conditions. The ff19SB force field was applied to standard amino acids in KRAS, GAFF (general AMBER force field) to MRTX1133, and TIP3P (intermolecular potential three point) to water molecules. Energy minimization was performed using the Amber software suite. The systems were then heated to 300 K, followed by NVT (constant volume and temperature) equilibration and NPT (constant pressure and temperature) production. Root mean square deviation (RMSD), root mean square fluctuation (RMSF), principal component analysis (PCA) and solvent-accessible surface area (SASA) of MRTX1133 and GDP were analyzed using cpptraj. The number of hydrogen bonds between regions and the dynamic cross-correlation matrix (DCCM) of amino acid movements were also calculated. Results ·RMSD analysis showed greater structural variation in KRAS in the KRASG12D/R68G system compared to the KRASG12D system. RMSF analysis revealed significantly higher fluctuations in the Switch Ⅰ and Switch Ⅱ regions of the KRASG12D/R68G system. PCA indicated that Switch Ⅰ and Switch Ⅱ in the KRASG12D/R68G system were more frequently in an open conformation. The distances between Switch Ⅰ and the P-loop, and between Switch Ⅱ and the P-loop, were larger in the KRASG12D/R68G system, indicating an expanded binding pocket for GDP and MRTX1133 compared to the KRASG12D system. SASA analysis indicated that both GDP and MRTX1133 had increased solvent exposure in the KRASG12D/R68G system. DCCM analysis revealed more decoupled movements among the Switch Ⅰ, Switch Ⅱ and P-loop regions in the KRASG12D/R68G system. Conclusion ·The KRASG12D/R68G mutation disrupts the interactions between the Switch Ⅰ and Switch Ⅱ regions, leading to their separation and the opening of the MRTX1133 binding pocket. This increases the solvent exposure of MRTX1133, accelerates its dissociation, and ultimately results in KRASG12D/R68G resistance to MRTX1133.
WANG Gaoming, CUI Ran, LI Yanjing, LIU Yingbin. Study on the mechanism of KRAS R68G secondary mutation-induced resistance to KRASG12D-targeted inhibitor MRTX1133. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2025, 45(6): 705-716 doi:10.3969/j.issn.1674-8115.2025.06.005
KRAS(Kirsten rat sarcoma viral oncogene homolog)蛋白是一种小GTP水解酶,属于RAS蛋白家族,在细胞信号转导过程中发挥重要作用,参与调控细胞的增殖、分化和凋亡等生理过程[1]。KRAS蛋白在结构上可分为核苷酸结合域(G域)和C末端高变区疏水尾[2]。核苷酸结合域由第1~165位氨基酸构成,包含P-loop(第10~17位氨基酸)、Switch Ⅰ(第32~38位氨基酸)和Switch Ⅱ(第59~76位氨基酸)等关键区域,负责GTP/GDP的结合和水解,调控下游的信号转导[2-3]。C末端疏水尾由第166位至最后一位氨基酸构成,负责将KRAS锚定到细胞膜上[2]。在生理条件下,KRAS在与GTP结合和与GDP结合的状态之间循环,发挥分子开关的作用。KRAS与GTP结合会激活下游的RAF/MEK/ERK和PI3K/AKT/mTOR等信号通路[4]。在GTP酶激活蛋白的帮助下,KRAS的GTP水解酶活性提高,将GTP水解为GDP,从而失去激活下游信号通路的功能[5]。鸟嘌呤核苷酸交换因子通过催化KRAS的GDP与GTP交换,使KRAS重新与GTP结合,促进下游信号通路的激活[6]。这种KRAS-GTP和KRAS-GDP状态之间的循环有序调控着细胞内信号的转导,从而调节细胞的正常生理活动。
从结构生物信息学研究合作组织(Research Collaboratory for Structural Bioinformatics,RCSB)蛋白质数据库(Protein Data Bank,PDB)中获取KRASG12D与MRTX1133相互作用的复合物的晶体结构数据(PDB ID: 7RPZ)[15],作为G12D体系进行分子动力学模拟的初始结构。使用PyMOL软件将G12D体系中KRAS的第68位精氨酸突变为甘氨酸,作为G12D_R68G体系进行分子动力学模拟的初始结构。由于7RPZ晶体结构中缺失了KRAS蛋白第2位氨基酸的坐标,因此对KRAS蛋白的模拟从第3位的谷氨酸开始至第169位的赖氨酸结束。
使用Amber22软件和AmberTools23软件[20]中的antechamber程序和键电荷校正(bond charge correction,BCC)的方法计算和构建MRTX1133的力场参数。使用LEaP程序构建和描绘KRAS-MRTX1133复合物的拓扑学参数和位置坐标。在LEaP程序中,依次载入KRASG12D/KRASG12D/R68G蛋白、GDP、MRTX1133和二价镁离子的初始坐标,形成蛋白-GDP-配体-离子的复合物。应用ff19SB力场[21]计算KRAS中标准氨基酸的力场参数,应用GAFF(general AMBER force field)力场[22]计算MRTX1133的力场参数,应用TIP3P(intermolecular potential three point)力场计算水分子的力场参数,GDP和二价镁离子的力场参数从Amber参数数据库中获取(http://amber.manchester.ac.uk/)。为模拟正常的生理盐水环境,将复合物溶解于正八面体的水盒子中,并加入Na+和Cl-对体系中的电荷进行中和,计算整个系统的拓扑学参数,完成G12D和G12D_R68G体系的构建。
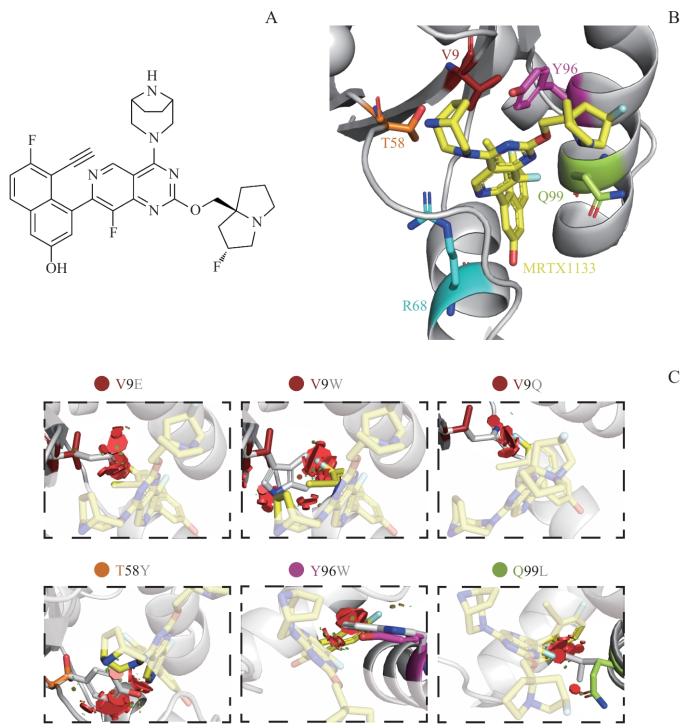
Note: A. Chemical structure of MRTX1133. B. The MRTX1133 binding pocket (PDB ID: 7RPZ). C. Six secondary mutations, including V9E, V9W, V9Q, T58Y, Y96W and Q99L, cause significant steric hindrance or block the formation of interactions between MRTX1133 and KRAS. Red areas stand for steric clashes.
Fig 1
MRTX1133 and its binding pocket in KRAS, and associated resistance mutations
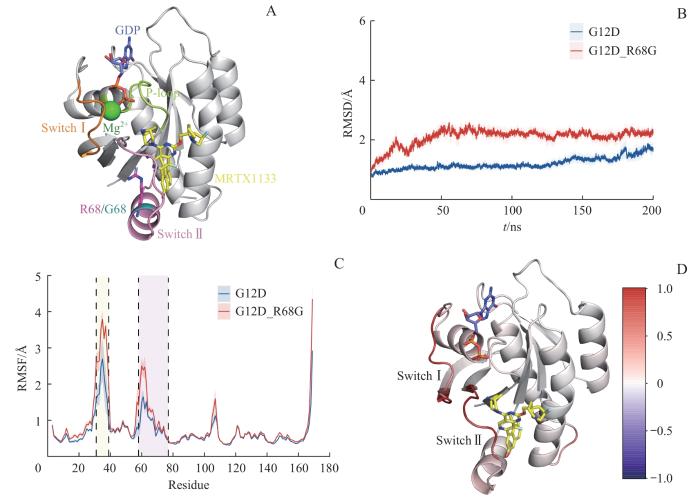
Note: A. The initial structures of the KRASG12D and KRASG12D/R68G systems. B. RMSDs of Cα atoms in KRASG12D and KRASG12D/R68G systems. C. RMSFs of each residue in KRASG12D and KRASG12D/R68G systems. Yellow region stands for the Switch Ⅰ region, while pink region represents the Switch Ⅱ region. D. The distribution of delta RMSF values of the KRASG12D/R68G system relative to the KRASG12D system. Red regions represent higher RMSF values, whereas blue regions stand for lower RMSF values.
Fig 2
Conformational dynamics of KRASG12D and KRASG12D/R68G
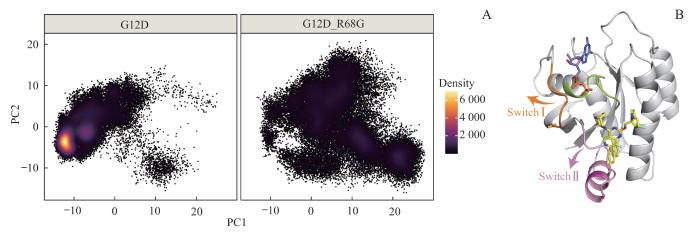
为了描绘2个体系全局的构象转变,对整个运动过程的轨迹进行PCA。根据每帧构象在前2个主成分(PC1和PC2)上的投影,绘制G12D和G12D_R68G体系的点密度图(图3A)。结果显示,与G12D体系相比,G12D_R68G体系具有明显较高的PC1值[(7.69±8.46) vs (-7.69±5.52),P<0.001,图3A]。将运动轨迹按照PC1值排序,得到KRAS蛋白随着PC1值增加而产生的构象变化(图3B),即Switch Ⅰ区域和Switch Ⅱ区域的柔性部分均远离P-loop区域而向外运动。综上所述,KRAS R68G继发突变破坏了KRASG12D的运动模式,促进KRAS向更加开放的构象转变。
Note: A. Projections of the first and second principal components (PC1 & PC2) from molecular dynamics simulations of KRASG12D and KRASG12D/R68G systems. B. Conformational changes along PC1.
Fig 3
Global conformational changes of the KRAS protein
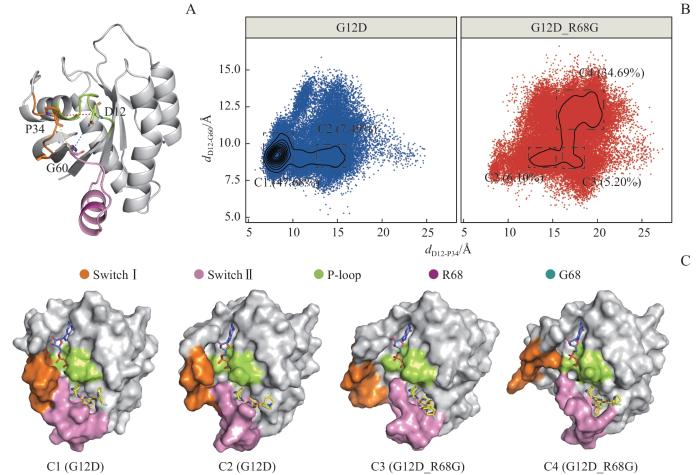
Note: A. Characteristic vectors indicating distances of Cα atoms between D12 and P34 (dD12-P34, red dotted line), between D12 and G60 (dD12-G60, blue dotted line), and between P34 and G60 (dP34-G60, yellow dotted line). B. Conformational landscapes generated using the dD12-P34 and dD12-G60 in KRASG12D (blue) and KRASG12D/R68G (red)systems. C. Surface representations of the representative structures in conformational clusters C1 to C4.
Fig 4
Conformational landscape and representative conformations of the KRAS protein
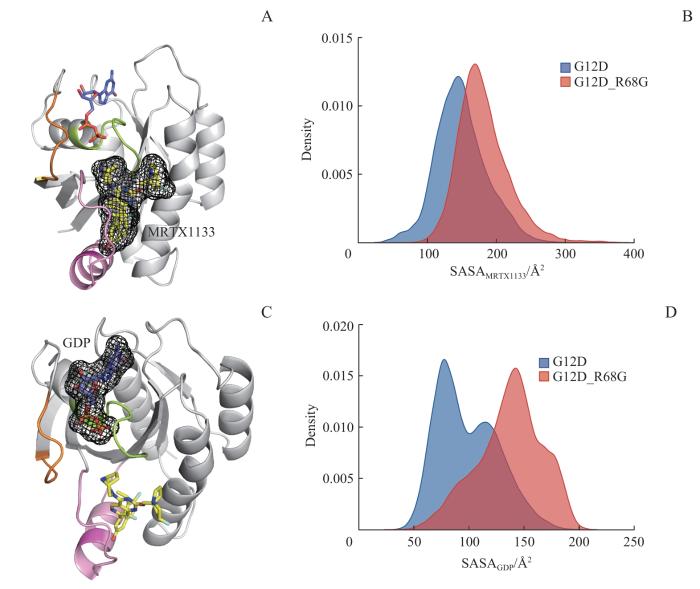
Note: A. Diagram of the SASA of MRTX1133. B. Density plots of SASA of MRTX1133 in KRASG12D and KRASG12D/R68G systems. C. Diagram of the SASA of GDP. D. Density plots of SASA of GDP in KRASG12D and KRASG12D/R68G systems.
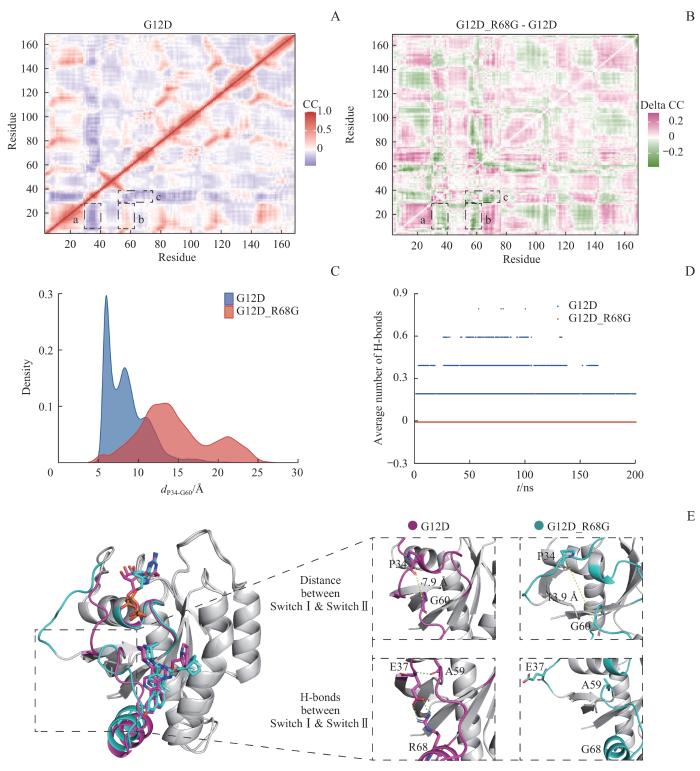
Note: A. DCCM plot of the KRASG12D system. B. Delta DCCM plot of the KRASG12D/R68G system relative to the KRASG12D system. Region a represents the decoupled motion between Switch Ⅰ and P-loop; region b indicates the decoupled motion between Switch Ⅱ and P-loop; region c suggests the decoupled motion between Switch Ⅰ and Switch Ⅱ. C. Distance distribution of Cα atoms between P34 and G60 (dP34-G60). D. Average hydrogen bonds formed between Switch Ⅰ and Switch Ⅱ during the simulations. E. Representative structures of KRASG12D (magenta) and KRASG12D/R68G (cyan) systems show the distance (yellow dotted line) and polar contacts (green dotted line) between Switch Ⅰ and Switch Ⅱ.
Fig 6
Dynamic relationship between Switch Ⅰ and Switch Ⅱ regions
为了更好地描绘Switch Ⅰ和Switch Ⅱ的位置关系,选取特征向量dP34-G60代表Switch Ⅰ和Switch Ⅱ之间的距离(图4A)。在整个运动过程中,G12D_R68G体系Switch Ⅰ和Switch Ⅱ之间的距离显著高于G12D体系[(14.50±4.52)Å vs (8.50±2.55)Å,P<0.001,图6C]。
WANG Gaoming and LI Yanjing designed the study. WANG Gaoming and CUI Ran performed molecular dynamics simulations and statistical analyses. WANG Gaoming wrote the manuscript. WANG Gaoming, LI Yanjing, and LIU Yingbin conducted an extensive literature review and discussed potential resistance mechanisms of KRASG12D to MRTX1133. LI Yanjing and LIU Yingbin revised the manuscript. All authors have read the final version of the paper and consented to its submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
HILLIG R C, SAUTIER B, SCHROEDER J, et al. Discovery of potent SOS1 inhibitors that block RAS activation via disruption of the RAS-SOS1 interaction[J]. Proc Natl Acad Sci USA, 2019, 116(7): 2551-2560.
WOOD K, HENSING T, MALIK R, et al. Prognostic and predictive value in KRAS in non-small-cell lung cancer: a review[J]. JAMA Oncol, 2016, 2(6): 805-812.
HUNTER J C, MANANDHAR A, CARRASCO M A, et al. Biochemical and structural analysis of common cancer-associated KRAS mutations[J]. Mol Cancer Res, 2015, 13(9): 1325-1335.
Cancer Genome Atlas Research Network. Integrated genomic characterization of pancreatic ductal adenocarcinoma[J]. Cancer Cell, 2017, 32(2): 185-203.e13.
LI M L, ZHANG Z, LI X G, et al. Whole-exome and targeted gene sequencing of gallbladder carcinoma identifies recurrent mutations in the ErbB pathway[J]. Nat Genet, 2014, 46(8): 872-876.
ROS J, VAGHI C, BARAIBAR I, et al. Targeting KRAS G12C mutation in colorectal cancer, a review: new arrows in the quiver[J]. Int J Mol Sci, 2024, 25(6): 3304.
WANG X L, ALLEN S, BLAKE J F, et al. Identification of MRTX1133, a noncovalent, potent, and selective KRASG12D inhibitor[J]. J Med Chem, 2022, 65(4): 3123-3133.
MAHADEVAN K K, MCANDREWS K M, LEBLEU V S, et al. KRASG12D inhibition reprograms the microenvironment of early and advanced pancreatic cancer to promote FAS-mediated killing by CD8+ T cells[J]. Cancer Cell, 2023, 41(9): 1606-1620.e8.
TANAKA N, LIN J J, LI C D, et al. Clinical acquired resistance to KRASG12C inhibition through a novel KRAS switch-Ⅱ pocket mutation and polyclonal alterations converging on RAS-MAPK reactivation[J]. Cancer Discov, 2021, 11(8): 1913-1922.
CHOI J, SHIN J Y, KIM T K, et al. Site-specific mutagenesis screening in KRASG12D mutant library to uncover resistance mechanisms to KRASG12D inhibitors[J]. Cancer Lett, 2024, 591: 216904.
ROE D R, CHEATHAM T E 3rd. PTRAJ and CPPTRAJ: software for processing and analysis of molecular dynamics trajectory data[J]. J Chem Theory Comput, 2013, 9(7): 3084-3095.
TIAN C, KASAVAJHALA K, BELFON K A A, et al. ff19SB: amino-acid-specific protein backbone parameters trained against quantum mechanics energy surfaces in solution[J]. J Chem Theory Comput, 2020, 16(1): 528-552.
YORK D M, WLODAWER A, PEDERSEN L G, et al. Atomic-level accuracy in simulations of large protein crystals[J]. Proc Natl Acad Sci USA, 1994, 91(18): 8715-8718.
WANG Y H, JI D, LEI C Y, et al. Mechanistic insights into the effect of phosphorylation on Ras conformational dynamics and its interactions with cell signaling proteins[J]. Comput Struct Biotechnol J, 2021, 19: 1184-1199.
ZHUANG H M, FAN J G, LI M Y, et al. Mechanistic insights into the clinical Y96D mutation with acquired resistance to AMG510 in the KRASG12C[J]. Front Oncol, 2022, 12: 915512.
OSTREM J M, PETERS U, SOS M L, et al. K-Ras(G12C) inhibitors allosterically control GTP affinity and effector interactions[J]. Nature, 2013, 503(7477): 548-551.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}