SIRT6, a member of the sirtuin family of histone deacetylases, belongs to the class Ⅲ longevity proteins and exhibits NAD+-dependent deacetylase and mono-ADP-ribosyltransferase activities. SIRT6 is primarily located in the cell nucleus and plays a pivotal role in regulating genomic stability and relative gene expression, participating in the control of key processes such as energy metabolism and aging. Given its crucial role in maintaining cellular homeostasis and organismal health, SIRT6 has emerged as a potential therapeutic target, sparking significant research interest in the development of targeted modulators. Activating the longevity protein with drugs may provide therapeutic strategies for age-associated diseases, including aging, metabolic syndrome, inflammation, and reproductive health issues. The review elaborates the structural characteristics, enzymatic activities, and biological functions of SIRT6, as well as the mechanisms of action, pharmacological activities, and clinical applications of various SIRT6 activators.
LIU Yonghui, TANG Li, LIANG Taigang, ZHANG Jian, FENG Li. Research progress in the role of SIRT6 in aging and metabolism. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2024, 44(11): 1439-1446 doi:10.3969/j.issn.1674-8115.2024.11.011
组蛋白赖氨酸的乙酰化是一种由组蛋白乙酰转移酶催化的翻译后修饰,而去乙酰化反应则由组蛋白去乙酰化酶(histone deacetylase,HDAC)介导。根据进化分析以及与酵母中不同去乙酰化酶的序列相似性,HDAC可分为4类:Ⅰ类(HDAC 1~3、8)、Ⅱ类(HDAC 4~7、9、10)、Ⅲ类(sirtuins)和Ⅳ类(HDAC 11)[1]。Sirtuins是一类与酿酒酵母中的沉默信息调节蛋白SIR2(silent information regulator 2)同源且高度保守的蛋白。Sirtuins有依赖于烟酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide,NAD+)的脱酰基酶活性和单ADP核糖基转移酶活性。哺乳动物sirtuins蛋白家族中包含7种同工酶(SIRT1~7)[2]。尽管sirtuins结构中含有NAD+结合位点且共享一个约包含275个残基的高度保守催化位点,但它们的N端延伸和C端延伸区域的大小和序列存在差异,这直接决定了其在细胞定位、催化酶活性、底物选择性和生物功能上的区别[3]。SIRT1、SIRT6和SIRT7主要定位于细胞核,参与调控基因稳定性和转录;SIRT2主要存在于胞质,可去乙酰化微管蛋白等胞质蛋白的乙酰化修饰;SIRT3~5主要位于线粒体,调控线粒体能量代谢相关通路[4]。研究[5]发现棕榈酸结合SIRT6后可使其穿梭到胞质中,催化长链脂酰辅酶A合成酶5(acyl-CoA synthetase long-chain family member 5,ACSL5)的去乙酰化,从而调控脂肪酸的氧化。近年来,关于SIRT6的结构特征、催化酶活性及相应的生物学功能也逐渐被阐明。
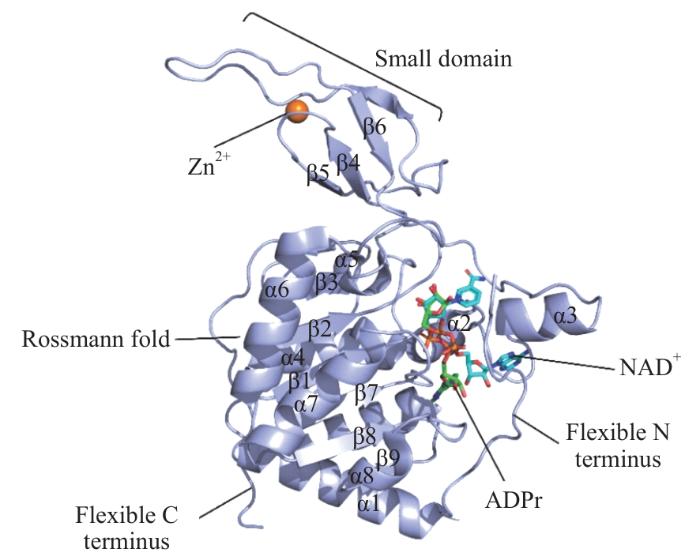
Note: The structural composition of SIRT6 is basically divided into Rossmann fold structure, small structural domains (including the zinc finger module), C-terminal extension (CTE) and N-terminal extension (NTE). SIRT6 is shown as a purple cartoon structure with lake blue stick structures for NAD+ and green for ADP ribose.
Fig 1
Secondary structure of the human protein SIRT6
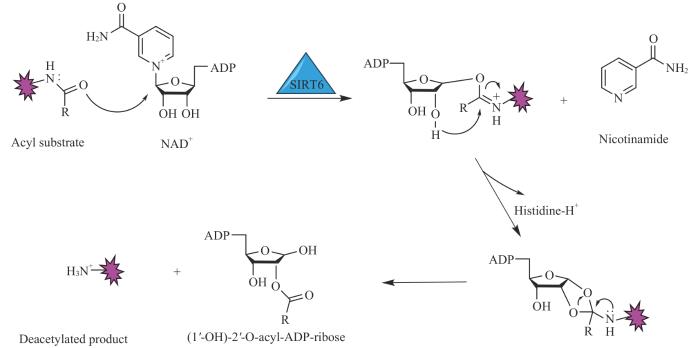
Note: NAD+ acts as a cofactor to assist SIRT6 in removing acylation groups from substrate lysine residues, resulting in the generation of NAM, ADP ribose, and the deacetylated product.
Fig 2
Mechanism of SIRT6 catalyzing deacylation
SIRT6体外体系中的去乙酰活性非常弱,催化速率大约只有其他sirtuins的千分之一。然而,它对包含DNA的完整核小体结构的乙酰化组蛋白具有很高的催化活性,这表明其高催化效率可能需要完整的生理环境中的染色质结构[11]。已知的SIRT6去乙酰化作用的底物有组蛋白和非组蛋白类型:组蛋白底物包括H3K9、H3K56、H3K18等[10];非组蛋白底物有PARP和组蛋白乙酰转移酶GCN5(general control of amino acid synthesis protein 5)等[18]。SIRT6的去乙酰化活性在多种生理过程中发挥着重要作用,例如代谢、衰老、炎症等。
3 SIRT6的生物学功能
3.1 衰老
自从sirtuins被证明是长寿蛋白后,它们就备受关注。衰老是一个复杂的生物过程,涉及多个维度,包括生理功能和健康水平的下降、虚弱感增加及寿命缩短[3]。研究[19]表明,SIRT6全身性缺失会导致小鼠出现典型的早衰综合征,表现为皮下脂肪流失、基因组不稳定性、致死性低血糖等症状,断奶后即死亡。这首次揭示了SIRT6在调节哺乳动物寿命方面的重要作用。研究[20]发现,SIRT6的过表达可以改善小鼠能量稳态并显著延长其寿命。事实上,SIRT6的寿命调控作用是一个极其复杂的过程,其中经历了多种生物过程的共同调控,如维持基因组稳定性和代谢稳态。越来越多的证据表明,DNA修复效率低下和DNA修复突变体是导致基因组不稳定性的主要原因,并最终加速衰老表型。相比之下,长寿物种的非同源末端连接(non-homologous end-joining,NHEJ)和同源重组(homologous recombination,HR)对双链断裂修复(double-strand break,DSB)的修复效率比短寿命物种更高[21]。令人更惊讶的是,在对18种不同寿命的啮齿动物进行比对后发现,SIRT6促进DSB修复的能力与寿命直接相关,SIRT6活性越高,则寿命越长[21]。SIRT6作为HR和NHEJ的上游调控因子,可以同时激活HR和NHEJ,从而提高修复效率和准确性[22]。另外,SIRT6还可以通过去乙酰化H3K9ac和H3K56ac实现端粒的正确复制。SIRT6的敲除导致端粒结构异常以及随机复制相关端粒序列丢失,最终导致基因组的不稳定性和细胞过早衰老[23]。能量限制可以通过抑制雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)-胰岛素样生长因子1(insulin-like growth factor 1,IGF1)通路,以及激活AMPK和sirtuins相关通路来延长寿命。研究[24]发现,SIRT6全身敲除导致的早衰缺陷主要与丝氨酸/苏氨酸激酶1-胰岛素样生长因子1(serine/threonine kinase-insulin like growth factor 1,AKT-IGF1)信号转导通路的增强相关。另外,SIRT6过表达可以降低小鼠血清中的IGF1水平,通过增加肝脏糖异生基因的表达和促进NAD+从头合成来维持老年鼠的能量稳态,从而实现延长30%左右的寿命[25]。总而言之,SIRT6可通过调节DNA损伤修复、端粒活性以及能量代谢等多个生物过程来共同调控衰老。
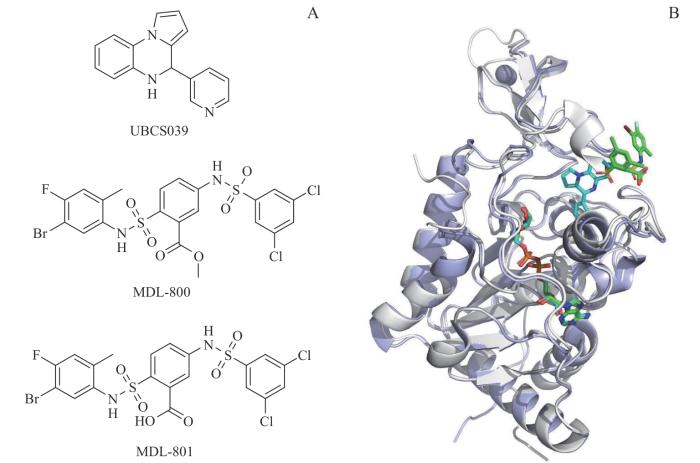
Note: A. Chemical structures of SIRT6 agonist, MDL-800, MDL-801, and UBCS039. B. The binding modes of MDL-801 or UBCS039 with SIRT6. MDL-801 and UBCS039 are shown in green and blue sticks, respectively.
Fig 3
Structure and binding modes of SIRT6 activators
LIU Yonghui drafted the original manuscript. TANG Li, LIANG Taigang, ZHANG Jian, and FENG Li participated in the reviewing and editing. All the authors have read the last version of paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
SHARMA A, MAHUR P, MUTHUKUMARAN J, et al. Shedding light on structure, function and regulation of human sirtuins: a comprehensive review[J]. 3 Biotech, 2023, 13(1): 29.
DZIDEK A, CZERWIŃSKA-LEDWIG O, ŻYCHOWSKA M, et al. The role of increased expression of sirtuin 6 in the prevention of premature aging pathomechanisms[J]. Int J Mol Sci, 2023, 24(11): 9655.
KLEIN M A, DENU J M. Biological and catalytic functions of sirtuin 6 as targets for small-molecule modulators[J]. J Biol Chem, 2020, 295(32): 11021-11041.
HOU T Y, TIAN Y, CAO Z Y, et al. Cytoplasmic SIRT6-mediated ACSL5 deacetylation impedes nonalcoholic fatty liver disease by facilitating hepatic fatty acid oxidation[J]. Mol Cell, 2022, 82(21): 4099-4115.e9.
MAHLKNECHT U, HO A D, VOELTER-MAHLKNECHT S. Chromosomal organization and fluorescence in situ hybridization of the human Sirtuin 6 gene[J]. Int J Oncol, 2006, 28(2): 447-456.
TENNEN R I, BERBER E, CHUA K F. Functional dissection of SIRT6: identification of domains that regulate histone deacetylase activity and chromatin localization[J]. Mech Ageing Dev, 2010, 131(3): 185-192.
WANG Z A, MARKERT J W, WHEDON S D, et al. Structural basis of sirtuin 6-catalyzed nucleosome deacetylation[J]. J Am Chem Soc, 2023, 145(12): 6811-6822.
KALOUS K S, WYNIA-SMITH S L, OLP M D, et al. Mechanism of Sirt1 NAD+-dependent protein deacetylase inhibition by cysteine S-nitrosation[J]. J Biol Chem, 2016, 291(49): 25398-25410.
RONNEBAUM S M, WU Y X, MCDONOUGH H, et al. The ubiquitin ligase CHIP prevents SirT6 degradation through noncanonical ubiquitination[J]. Mol Cell Biol, 2013, 33(22): 4461-4472.
JIN L, WEI W T, JIANG Y B, et al. Crystal structures of human SIRT3 displaying substrate-induced conformational changes[J]. J Biol Chem, 2009, 284(36): 24394-24405.
BARAN M, MIZIAK P, STEPULAK A, et al. The role of sirtuin 6 in the deacetylation of histone proteins as a factor in the progression of neoplastic disease[J]. Int J Mol Sci, 2023, 25(1): 497.
QIU B Q, LI S, LI M T, et al. KAT8 acetylation-controlled lipolysis affects the invasive and migratory potential of colorectal cancer cells[J]. Cell Death Dis, 2023, 14(2): 164.
MOSTOSLAVSKY R, CHUA K F, LOMBARD D B, et al. Genomic instability and aging-like phenotype in the absence of mammalian SIRT6[J]. Cell, 2006, 124(2): 315-329.
TIAN X, FIRSANOV D, ZHANG Z H, et al. SIRT6 is responsible for more efficient DNA double-strand break repair in long-lived species[J]. Cell, 2019, 177(3): 622-638.e22.
MICHISHITA E, MCCORD R A, BOXER L D, et al. Cell cycle-dependent deacetylation of telomeric histone H3 lysine K56 by human SIRT6[J]. Cell Cycle, 2009, 8(16): 2664-2666.
XIAO C Y, KIM H S, LAHUSEN T, et al. SIRT6 deficiency results in severe hypoglycemia by enhancing both basal and insulin-stimulated glucose uptake in mice[J]. J Biol Chem, 2010, 285(47): 36776-36784.
SIMON M, YANG J P, GIGAS J, et al. A rare human centenarian variant of SIRT6 enhances genome stability and interaction with Lamin A[J]. EMBO J, 2022, 41(21): e110393.
FERRER C M, ALDERS M, POSTMA A V, et al. An inactivating mutation in the histone deacetylase SIRT6 causes human perinatal lethality[J]. Genes Dev, 2018, 32(5/6): 373-388.
KIM H S, XIAO C Y, WANG R H, et al. Hepatic-specific disruption of SIRT6 in mice results in fatty liver formation due to enhanced glycolysis and triglyceride synthesis[J]. Cell Metab, 2010, 12(3): 224-236.
TAO R Y, XIONG X W, DEPINHO R A, et al. FoxO3 transcription factor and Sirt6 deacetylase regulate low density lipoprotein (LDL)-cholesterol homeostasis via control of the proprotein convertase subtilisin/kexin type 9 (Pcsk9) gene expression[J]. J Biol Chem, 2013, 288(41): 29252-29259.
DOMINY J E Jr, LEE Y, JEDRYCHOWSKI M P, et al. The deacetylase Sirt6 activates the acetyltransferase GCN5 and suppresses hepatic gluconeogenesis[J]. Mol Cell, 2012, 48(6): 900-913.
BIAN C, ZHANG R J, WANG Y X, et al. Sirtuin 6 affects glucose reabsorption and gluconeogenesis in type 1 diabetes via FoxO1[J]. Mol Cell Endocrinol, 2022, 547: 111597.
SUN H L, WU Y R, FU D J, et al. SIRT6 regulates osteogenic differentiation of rat bone marrow mesenchymal stem cells partially via suppressing the nuclear factor-κB signaling pathway[J]. Stem Cells, 2014, 32(7): 1943-1955.
LEE Y, KA S O, CHA H N, et al. Myeloid sirtuin 6 deficiency causes insulin resistance in high-fat diet-fed mice by eliciting macrophage polarization toward an M1 phenotype[J]. Diabetes, 2017, 66(10): 2659-2668.
DING Y N, WANG T T, LV S J, et al. SIRT6 is an epigenetic repressor of thoracic aortic aneurysms via inhibiting inflammation and senescence[J]. Signal Transduct Target Ther, 2023, 8(1): 255.
TATONE C, EMIDIO G D, BARBONETTI A, et al. Sirtuins in gamete biology and reproductive physiology: emerging roles and therapeutic potential in female and male infertility[J]. Hum Reprod Update, 2018, 24(3): 267-289.
HAN L S, GE J, ZHANG L, et al. Sirt6 depletion causes spindle defects and chromosome misalignment during meiosis of mouse oocyte[J]. Sci Rep, 2015, 5: 15366.
LIU W J, ZHANG X M, WANG N, et al. Calorie restriction inhibits ovarian follicle development and follicle loss through activating SIRT1 signaling in mice[J]. Eur J Med Res, 2015, 20(1): 22.
LI L Y, HUA R, HU K Q, et al. SIRT6 deficiency causes ovarian hypoplasia by affecting Plod1-related collagen formation[J]. Aging Cell, 2024, 23(2): e14031.
MICHISHITA E, PARK J Y, BURNESKIS J M, et al. Evolutionarily conserved and nonconserved cellular localizations and functions of human SIRT proteins[J]. Mol Biol Cell, 2005, 16(10): 4623-4635.
PASCOAL G F L, GERALDI M V, MARÓSTICA M R Jr, et al. Effect of paternal diet on spermatogenesis and offspring health: focus on epigenetics and interventions with food bioactive compounds[J]. Nutrients, 2022, 14(10): 2150.
WU Y F, YING J H, ZHU X Y, et al. Pachymic acid suppresses the inflammatory response of chondrocytes and alleviates the progression of osteoarthritis via regulating the Sirtuin 6/NF-κB signal axis[J]. Int Immunopharmacol, 2023, 124(Pt A): 110854.
LEE A, GU H, GWON M H, et al. Hesperetin suppresses LPS/high glucose-induced inflammatory responses via TLR/MyD88/NF-κB signaling pathways in THP-1 cells[J]. Nutr Res Pract, 2021, 15(5): 591-603.
PAN Z S, GUO J Y, TANG K J, et al. Ginsenoside Rc modulates SIRT6-NRF2 interaction to alleviate alcoholic liver disease[J]. J Agric Food Chem, 2022, 70(44): 14220-14234.
WU R Y, JIAN T, DING X Q, et al. Total sesquiterpene glycosides from loquat leaves ameliorate HFD-induced insulin resistance by modulating IRS-1/GLUT4, TRPV1, and SIRT6/Nrf2 signaling pathways[J]. Oxid Med Cell Longev, 2021, 2021: 4706410.
IACHETTINI S, TRISCIUOGLIO D, ROTILI D, et al. Pharmacological activation of SIRT6 triggers lethal autophagy in human cancer cells[J]. Cell Death Dis, 2018, 9(10): 996.
JIAO F Z, ZHANG Z W, HU H T, et al. SIRT6 activator UBCS039 inhibits thioacetamide-induced hepatic injury in vitro and in vivo[J]. Front Pharmacol, 2022, 13: 837544.
ZHANG J H, LI Y P, LIU Q H, et al. Sirt6 alleviated liver fibrosis by deacetylating conserved lysine 54 on Smad2 in hepatic stellate cells[J]. Hepatology, 2021, 73(3): 1140-1157.
CHEN Y, CHEN J Y, SUN X X, et al. The SIRT6 activator MDL-800 improves genomic stability and pluripotency of old murine-derived iPS cells[J]. Aging Cell, 2020, 19(8): e13185.
... SIRT6体外体系中的去乙酰活性非常弱,催化速率大约只有其他sirtuins的千分之一.然而,它对包含DNA的完整核小体结构的乙酰化组蛋白具有很高的催化活性,这表明其高催化效率可能需要完整的生理环境中的染色质结构[11].已知的SIRT6去乙酰化作用的底物有组蛋白和非组蛋白类型:组蛋白底物包括H3K9、H3K56、H3K18等[10];非组蛋白底物有PARP和组蛋白乙酰转移酶GCN5(general control of amino acid synthesis protein 5)等[18].SIRT6的去乙酰化活性在多种生理过程中发挥着重要作用,例如代谢、衰老、炎症等. ...
... SIRT6体外体系中的去乙酰活性非常弱,催化速率大约只有其他sirtuins的千分之一.然而,它对包含DNA的完整核小体结构的乙酰化组蛋白具有很高的催化活性,这表明其高催化效率可能需要完整的生理环境中的染色质结构[11].已知的SIRT6去乙酰化作用的底物有组蛋白和非组蛋白类型:组蛋白底物包括H3K9、H3K56、H3K18等[10];非组蛋白底物有PARP和组蛋白乙酰转移酶GCN5(general control of amino acid synthesis protein 5)等[18].SIRT6的去乙酰化活性在多种生理过程中发挥着重要作用,例如代谢、衰老、炎症等. ...
... SIRT6体外体系中的去乙酰活性非常弱,催化速率大约只有其他sirtuins的千分之一.然而,它对包含DNA的完整核小体结构的乙酰化组蛋白具有很高的催化活性,这表明其高催化效率可能需要完整的生理环境中的染色质结构[11].已知的SIRT6去乙酰化作用的底物有组蛋白和非组蛋白类型:组蛋白底物包括H3K9、H3K56、H3K18等[10];非组蛋白底物有PARP和组蛋白乙酰转移酶GCN5(general control of amino acid synthesis protein 5)等[18].SIRT6的去乙酰化活性在多种生理过程中发挥着重要作用,例如代谢、衰老、炎症等. ...
1
... 自从sirtuins被证明是长寿蛋白后,它们就备受关注.衰老是一个复杂的生物过程,涉及多个维度,包括生理功能和健康水平的下降、虚弱感增加及寿命缩短[3].研究[19]表明,SIRT6全身性缺失会导致小鼠出现典型的早衰综合征,表现为皮下脂肪流失、基因组不稳定性、致死性低血糖等症状,断奶后即死亡.这首次揭示了SIRT6在调节哺乳动物寿命方面的重要作用.研究[20]发现,SIRT6的过表达可以改善小鼠能量稳态并显著延长其寿命.事实上,SIRT6的寿命调控作用是一个极其复杂的过程,其中经历了多种生物过程的共同调控,如维持基因组稳定性和代谢稳态.越来越多的证据表明,DNA修复效率低下和DNA修复突变体是导致基因组不稳定性的主要原因,并最终加速衰老表型.相比之下,长寿物种的非同源末端连接(non-homologous end-joining,NHEJ)和同源重组(homologous recombination,HR)对双链断裂修复(double-strand break,DSB)的修复效率比短寿命物种更高[21].令人更惊讶的是,在对18种不同寿命的啮齿动物进行比对后发现,SIRT6促进DSB修复的能力与寿命直接相关,SIRT6活性越高,则寿命越长[21].SIRT6作为HR和NHEJ的上游调控因子,可以同时激活HR和NHEJ,从而提高修复效率和准确性[22].另外,SIRT6还可以通过去乙酰化H3K9ac和H3K56ac实现端粒的正确复制.SIRT6的敲除导致端粒结构异常以及随机复制相关端粒序列丢失,最终导致基因组的不稳定性和细胞过早衰老[23].能量限制可以通过抑制雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)-胰岛素样生长因子1(insulin-like growth factor 1,IGF1)通路,以及激活AMPK和sirtuins相关通路来延长寿命.研究[24]发现,SIRT6全身敲除导致的早衰缺陷主要与丝氨酸/苏氨酸激酶1-胰岛素样生长因子1(serine/threonine kinase-insulin like growth factor 1,AKT-IGF1)信号转导通路的增强相关.另外,SIRT6过表达可以降低小鼠血清中的IGF1水平,通过增加肝脏糖异生基因的表达和促进NAD+从头合成来维持老年鼠的能量稳态,从而实现延长30%左右的寿命[25].总而言之,SIRT6可通过调节DNA损伤修复、端粒活性以及能量代谢等多个生物过程来共同调控衰老. ...
1
... 自从sirtuins被证明是长寿蛋白后,它们就备受关注.衰老是一个复杂的生物过程,涉及多个维度,包括生理功能和健康水平的下降、虚弱感增加及寿命缩短[3].研究[19]表明,SIRT6全身性缺失会导致小鼠出现典型的早衰综合征,表现为皮下脂肪流失、基因组不稳定性、致死性低血糖等症状,断奶后即死亡.这首次揭示了SIRT6在调节哺乳动物寿命方面的重要作用.研究[20]发现,SIRT6的过表达可以改善小鼠能量稳态并显著延长其寿命.事实上,SIRT6的寿命调控作用是一个极其复杂的过程,其中经历了多种生物过程的共同调控,如维持基因组稳定性和代谢稳态.越来越多的证据表明,DNA修复效率低下和DNA修复突变体是导致基因组不稳定性的主要原因,并最终加速衰老表型.相比之下,长寿物种的非同源末端连接(non-homologous end-joining,NHEJ)和同源重组(homologous recombination,HR)对双链断裂修复(double-strand break,DSB)的修复效率比短寿命物种更高[21].令人更惊讶的是,在对18种不同寿命的啮齿动物进行比对后发现,SIRT6促进DSB修复的能力与寿命直接相关,SIRT6活性越高,则寿命越长[21].SIRT6作为HR和NHEJ的上游调控因子,可以同时激活HR和NHEJ,从而提高修复效率和准确性[22].另外,SIRT6还可以通过去乙酰化H3K9ac和H3K56ac实现端粒的正确复制.SIRT6的敲除导致端粒结构异常以及随机复制相关端粒序列丢失,最终导致基因组的不稳定性和细胞过早衰老[23].能量限制可以通过抑制雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)-胰岛素样生长因子1(insulin-like growth factor 1,IGF1)通路,以及激活AMPK和sirtuins相关通路来延长寿命.研究[24]发现,SIRT6全身敲除导致的早衰缺陷主要与丝氨酸/苏氨酸激酶1-胰岛素样生长因子1(serine/threonine kinase-insulin like growth factor 1,AKT-IGF1)信号转导通路的增强相关.另外,SIRT6过表达可以降低小鼠血清中的IGF1水平,通过增加肝脏糖异生基因的表达和促进NAD+从头合成来维持老年鼠的能量稳态,从而实现延长30%左右的寿命[25].总而言之,SIRT6可通过调节DNA损伤修复、端粒活性以及能量代谢等多个生物过程来共同调控衰老. ...
2
... 自从sirtuins被证明是长寿蛋白后,它们就备受关注.衰老是一个复杂的生物过程,涉及多个维度,包括生理功能和健康水平的下降、虚弱感增加及寿命缩短[3].研究[19]表明,SIRT6全身性缺失会导致小鼠出现典型的早衰综合征,表现为皮下脂肪流失、基因组不稳定性、致死性低血糖等症状,断奶后即死亡.这首次揭示了SIRT6在调节哺乳动物寿命方面的重要作用.研究[20]发现,SIRT6的过表达可以改善小鼠能量稳态并显著延长其寿命.事实上,SIRT6的寿命调控作用是一个极其复杂的过程,其中经历了多种生物过程的共同调控,如维持基因组稳定性和代谢稳态.越来越多的证据表明,DNA修复效率低下和DNA修复突变体是导致基因组不稳定性的主要原因,并最终加速衰老表型.相比之下,长寿物种的非同源末端连接(non-homologous end-joining,NHEJ)和同源重组(homologous recombination,HR)对双链断裂修复(double-strand break,DSB)的修复效率比短寿命物种更高[21].令人更惊讶的是,在对18种不同寿命的啮齿动物进行比对后发现,SIRT6促进DSB修复的能力与寿命直接相关,SIRT6活性越高,则寿命越长[21].SIRT6作为HR和NHEJ的上游调控因子,可以同时激活HR和NHEJ,从而提高修复效率和准确性[22].另外,SIRT6还可以通过去乙酰化H3K9ac和H3K56ac实现端粒的正确复制.SIRT6的敲除导致端粒结构异常以及随机复制相关端粒序列丢失,最终导致基因组的不稳定性和细胞过早衰老[23].能量限制可以通过抑制雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)-胰岛素样生长因子1(insulin-like growth factor 1,IGF1)通路,以及激活AMPK和sirtuins相关通路来延长寿命.研究[24]发现,SIRT6全身敲除导致的早衰缺陷主要与丝氨酸/苏氨酸激酶1-胰岛素样生长因子1(serine/threonine kinase-insulin like growth factor 1,AKT-IGF1)信号转导通路的增强相关.另外,SIRT6过表达可以降低小鼠血清中的IGF1水平,通过增加肝脏糖异生基因的表达和促进NAD+从头合成来维持老年鼠的能量稳态,从而实现延长30%左右的寿命[25].总而言之,SIRT6可通过调节DNA损伤修复、端粒活性以及能量代谢等多个生物过程来共同调控衰老. ...
... [21].SIRT6作为HR和NHEJ的上游调控因子,可以同时激活HR和NHEJ,从而提高修复效率和准确性[22].另外,SIRT6还可以通过去乙酰化H3K9ac和H3K56ac实现端粒的正确复制.SIRT6的敲除导致端粒结构异常以及随机复制相关端粒序列丢失,最终导致基因组的不稳定性和细胞过早衰老[23].能量限制可以通过抑制雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)-胰岛素样生长因子1(insulin-like growth factor 1,IGF1)通路,以及激活AMPK和sirtuins相关通路来延长寿命.研究[24]发现,SIRT6全身敲除导致的早衰缺陷主要与丝氨酸/苏氨酸激酶1-胰岛素样生长因子1(serine/threonine kinase-insulin like growth factor 1,AKT-IGF1)信号转导通路的增强相关.另外,SIRT6过表达可以降低小鼠血清中的IGF1水平,通过增加肝脏糖异生基因的表达和促进NAD+从头合成来维持老年鼠的能量稳态,从而实现延长30%左右的寿命[25].总而言之,SIRT6可通过调节DNA损伤修复、端粒活性以及能量代谢等多个生物过程来共同调控衰老. ...
1
... 自从sirtuins被证明是长寿蛋白后,它们就备受关注.衰老是一个复杂的生物过程,涉及多个维度,包括生理功能和健康水平的下降、虚弱感增加及寿命缩短[3].研究[19]表明,SIRT6全身性缺失会导致小鼠出现典型的早衰综合征,表现为皮下脂肪流失、基因组不稳定性、致死性低血糖等症状,断奶后即死亡.这首次揭示了SIRT6在调节哺乳动物寿命方面的重要作用.研究[20]发现,SIRT6的过表达可以改善小鼠能量稳态并显著延长其寿命.事实上,SIRT6的寿命调控作用是一个极其复杂的过程,其中经历了多种生物过程的共同调控,如维持基因组稳定性和代谢稳态.越来越多的证据表明,DNA修复效率低下和DNA修复突变体是导致基因组不稳定性的主要原因,并最终加速衰老表型.相比之下,长寿物种的非同源末端连接(non-homologous end-joining,NHEJ)和同源重组(homologous recombination,HR)对双链断裂修复(double-strand break,DSB)的修复效率比短寿命物种更高[21].令人更惊讶的是,在对18种不同寿命的啮齿动物进行比对后发现,SIRT6促进DSB修复的能力与寿命直接相关,SIRT6活性越高,则寿命越长[21].SIRT6作为HR和NHEJ的上游调控因子,可以同时激活HR和NHEJ,从而提高修复效率和准确性[22].另外,SIRT6还可以通过去乙酰化H3K9ac和H3K56ac实现端粒的正确复制.SIRT6的敲除导致端粒结构异常以及随机复制相关端粒序列丢失,最终导致基因组的不稳定性和细胞过早衰老[23].能量限制可以通过抑制雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)-胰岛素样生长因子1(insulin-like growth factor 1,IGF1)通路,以及激活AMPK和sirtuins相关通路来延长寿命.研究[24]发现,SIRT6全身敲除导致的早衰缺陷主要与丝氨酸/苏氨酸激酶1-胰岛素样生长因子1(serine/threonine kinase-insulin like growth factor 1,AKT-IGF1)信号转导通路的增强相关.另外,SIRT6过表达可以降低小鼠血清中的IGF1水平,通过增加肝脏糖异生基因的表达和促进NAD+从头合成来维持老年鼠的能量稳态,从而实现延长30%左右的寿命[25].总而言之,SIRT6可通过调节DNA损伤修复、端粒活性以及能量代谢等多个生物过程来共同调控衰老. ...
1
... 自从sirtuins被证明是长寿蛋白后,它们就备受关注.衰老是一个复杂的生物过程,涉及多个维度,包括生理功能和健康水平的下降、虚弱感增加及寿命缩短[3].研究[19]表明,SIRT6全身性缺失会导致小鼠出现典型的早衰综合征,表现为皮下脂肪流失、基因组不稳定性、致死性低血糖等症状,断奶后即死亡.这首次揭示了SIRT6在调节哺乳动物寿命方面的重要作用.研究[20]发现,SIRT6的过表达可以改善小鼠能量稳态并显著延长其寿命.事实上,SIRT6的寿命调控作用是一个极其复杂的过程,其中经历了多种生物过程的共同调控,如维持基因组稳定性和代谢稳态.越来越多的证据表明,DNA修复效率低下和DNA修复突变体是导致基因组不稳定性的主要原因,并最终加速衰老表型.相比之下,长寿物种的非同源末端连接(non-homologous end-joining,NHEJ)和同源重组(homologous recombination,HR)对双链断裂修复(double-strand break,DSB)的修复效率比短寿命物种更高[21].令人更惊讶的是,在对18种不同寿命的啮齿动物进行比对后发现,SIRT6促进DSB修复的能力与寿命直接相关,SIRT6活性越高,则寿命越长[21].SIRT6作为HR和NHEJ的上游调控因子,可以同时激活HR和NHEJ,从而提高修复效率和准确性[22].另外,SIRT6还可以通过去乙酰化H3K9ac和H3K56ac实现端粒的正确复制.SIRT6的敲除导致端粒结构异常以及随机复制相关端粒序列丢失,最终导致基因组的不稳定性和细胞过早衰老[23].能量限制可以通过抑制雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)-胰岛素样生长因子1(insulin-like growth factor 1,IGF1)通路,以及激活AMPK和sirtuins相关通路来延长寿命.研究[24]发现,SIRT6全身敲除导致的早衰缺陷主要与丝氨酸/苏氨酸激酶1-胰岛素样生长因子1(serine/threonine kinase-insulin like growth factor 1,AKT-IGF1)信号转导通路的增强相关.另外,SIRT6过表达可以降低小鼠血清中的IGF1水平,通过增加肝脏糖异生基因的表达和促进NAD+从头合成来维持老年鼠的能量稳态,从而实现延长30%左右的寿命[25].总而言之,SIRT6可通过调节DNA损伤修复、端粒活性以及能量代谢等多个生物过程来共同调控衰老. ...
2
... 自从sirtuins被证明是长寿蛋白后,它们就备受关注.衰老是一个复杂的生物过程,涉及多个维度,包括生理功能和健康水平的下降、虚弱感增加及寿命缩短[3].研究[19]表明,SIRT6全身性缺失会导致小鼠出现典型的早衰综合征,表现为皮下脂肪流失、基因组不稳定性、致死性低血糖等症状,断奶后即死亡.这首次揭示了SIRT6在调节哺乳动物寿命方面的重要作用.研究[20]发现,SIRT6的过表达可以改善小鼠能量稳态并显著延长其寿命.事实上,SIRT6的寿命调控作用是一个极其复杂的过程,其中经历了多种生物过程的共同调控,如维持基因组稳定性和代谢稳态.越来越多的证据表明,DNA修复效率低下和DNA修复突变体是导致基因组不稳定性的主要原因,并最终加速衰老表型.相比之下,长寿物种的非同源末端连接(non-homologous end-joining,NHEJ)和同源重组(homologous recombination,HR)对双链断裂修复(double-strand break,DSB)的修复效率比短寿命物种更高[21].令人更惊讶的是,在对18种不同寿命的啮齿动物进行比对后发现,SIRT6促进DSB修复的能力与寿命直接相关,SIRT6活性越高,则寿命越长[21].SIRT6作为HR和NHEJ的上游调控因子,可以同时激活HR和NHEJ,从而提高修复效率和准确性[22].另外,SIRT6还可以通过去乙酰化H3K9ac和H3K56ac实现端粒的正确复制.SIRT6的敲除导致端粒结构异常以及随机复制相关端粒序列丢失,最终导致基因组的不稳定性和细胞过早衰老[23].能量限制可以通过抑制雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)-胰岛素样生长因子1(insulin-like growth factor 1,IGF1)通路,以及激活AMPK和sirtuins相关通路来延长寿命.研究[24]发现,SIRT6全身敲除导致的早衰缺陷主要与丝氨酸/苏氨酸激酶1-胰岛素样生长因子1(serine/threonine kinase-insulin like growth factor 1,AKT-IGF1)信号转导通路的增强相关.另外,SIRT6过表达可以降低小鼠血清中的IGF1水平,通过增加肝脏糖异生基因的表达和促进NAD+从头合成来维持老年鼠的能量稳态,从而实现延长30%左右的寿命[25].总而言之,SIRT6可通过调节DNA损伤修复、端粒活性以及能量代谢等多个生物过程来共同调控衰老. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}