Objective ·To prepare a serine protease inhibitor (Serpin) derived from Tannerella which is associated with periodontosis, and analyze its specificity in inhibiting target proteases and its structural characteristics. Methods ·Through amino acid sequence analysis, a Serpin from the human oral microbiome database (eHOMD) was selected and expressed in Escherichia coli. The recombinant protein was purified using methods such as nickel ion affinity chromatography. Its specificity in inhibiting serine proteases was analyzed, followed by an analysis of its three-dimensional spatial structure using structural biology methods. Results ·A novel Serpin, named Tannerpin-M, with methionine as the active center P1 residue, was identified, and a high-purity recombinant protein was successfully prepared from Escherichia coli BL21 (DE3). Further activity testing demonstrated that recombinant Tannerpin-M could effectively form SDS-stable covalent complexes with proteases derived from granulocytes (human neutrophil elastase, cathepsin G, and proteinase 3), as well as with other proteases including kallikrein 1 (KLK1), KLK7, and elastase. Tannerpin-M inhibited KLK7 with a second-order association rate constant of 4.12×104 L/(mol·s). The crystal structure of Tannerpin-M in its relaxed state conformation was resolved at a resolution of 2.4 Å (1 Å=0.1 nm). It revealed that Tannerpin-M possessed a significantly elongated reactive center loop and could undergo the classical conformational transition from a stressed to a relaxed state. Conclusion ·Tannerpin-M, derived from oral pathogenic bacteria, is a typical inhibitory Serpin, and can effectively inhibit the serine protease released by granulocytes, by which it may protect the oral pathogenic bacteria from attacks of the human immune system.
PAN Zihao, XU Jiawei, ZHOU Aiwu. Inhibition of Tannerpin-M encoded by periodontal pathogens on serine proteases released by granulocytes. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2024, 44(12): 1536-1544 doi:10.3969/j.issn.1674-8115.2024.12.006
由于Serpin家族蛋白广泛存在于动植物以及某些微生物中,均由350~400个氨基酸组成,序列同源性往往大于25%[4-6]。典型的Serpin通常包括3个主要的β折叠(A、B、C)、8~9个α螺旋(hA~hI)和1个暴露在分子表面的中心环(reactive center loop,RCL)[7]。蛋白酶可以识别并切割RCL中特异的氨基酸(P1-P1' 残基)间的肽键,诱使Serpin发生一种独特的从紧张态到松弛态(stressed-to-relaxed,S-to-R)转换的构象变化,并且与蛋白酶形成一种对SDS稳定的共价相连的复合物,从而使蛋白酶失活[8-9]。大量研究表明,Serpin家族成员在生物体中可以通过抑制蛋白酶活性调控重要的生理过程,如血液凝固[10]、免疫反应[11]、细胞死亡等[12-13]。因此,我们推测这些牙周病原菌也能通过表达和分泌一些Serpin抑制宿主的蛋白酶,保护自身免受宿主免疫系统的杀伤,帮助病原体逃避免疫监控。
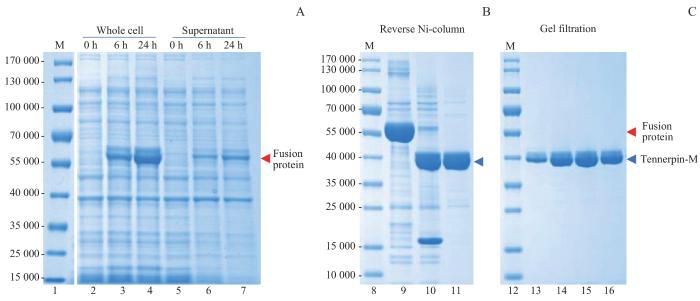
Note: A. Protein expression of Tannerpin-M fusion protein in whole bacterial cell lysate and lysis supernatant. M—marker (Lane 1,8 and 12); Lane 2‒4—whole cell lysate of Tannerpin-M expression upon IPTG induction at 0, 6 and 24 h; Lane 5‒7—supernatant of Tannerpin-M expression cells upon IPTG induction at 0, 6 and 24 h respectively. B. Enzyme digestion of the fusion protein and purification by reverse nickel ion affinity column (Ni-column). Lane 9—HisTrap-purified SUMO3-Tannerpin-M fusion protein; Lane 10—fusion protein digested with SENP2; Lane 11—flow through of fusion protein digestion from a HisTrap column. C. Gel filtration purification of Tannerpin-M. Lane13‒16—fractions of Tannerpin-M from a gel filtration column.
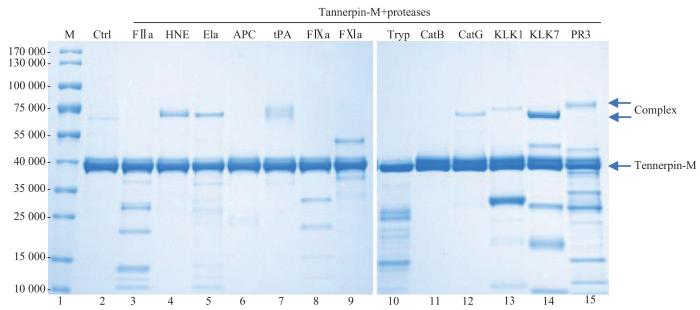
Note: M—marker (Lane 1); Lane 2—Tannerpin-M control (Ctrl). The other lanes are mixed solutions of Tannerpin-M with different serine proteases; Lane 3—FⅡa; Lane 4—HNE; Lane 5—elastase; Lane 6—APC; Lane 7—t-PA; Lane 8—FⅨa; Lane 9—FⅪa; Lane 10—trypsin; Lane 11—CatB; Lane 12—CatG; Lane 13—KLK1; Lane 14—KLK7; Lane 15—PR3.
Fig 3
Inhibition specificity of Tannerpin-M towards serine proteases
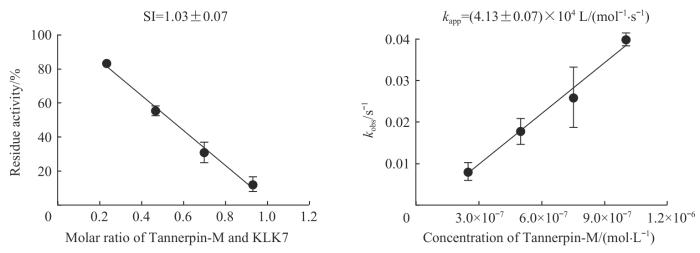
Note: The SI value of Tannerpin-M in inhibiting KLK7 was determined by mixing different ratios of Serpin and protease, with the remaining protease activity plotted against the ratio (left). kapp value of Tannerpin-M in inhibiting KLK7 was obtained by plotting the kobs values against the concentrations of Tannerpin-M (right).
Fig 4
Kinetic measurement of the inhibition of Tannerpin-M on KLK7
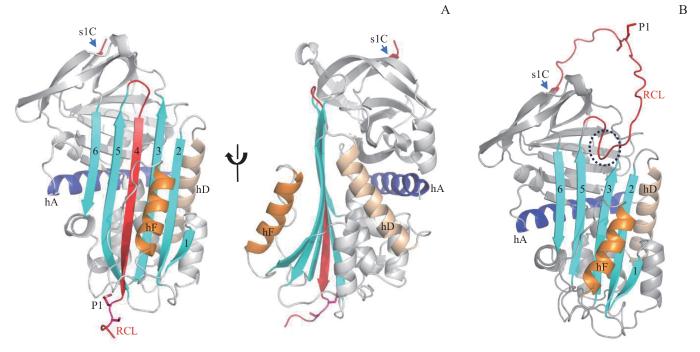
Note: A. Crystal structure of cleaved Tannerpin-M. B. Predicted structure of native Tannerpin-M. The central β-sheet A is colored in cyan with the strands traditionally numbered 1‒6 from right to left. RCL is colored in red, helix A (hA) is colored in purple, helix F (hF) is colored in orange, and helix D (hD) is colored in gray. P1-P1' residue is shown in sticks.
ZHOU Aiwu designed this study and supervised writing of the paper. PAN Zihao and XU Jiawei completed all the experiments. XU Jiawei wrote and revised the manuscript. All the authors have read the last version of the paper and consented for submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
LEYS E J, LYONS S R, MOESCHBERGER M L, et al. Association of Bacteroides forsythus and a novel Bacteroides phylotype with periodontitis[J]. J Clin Microbiol, 2002, 40(3): 821-825.
FARRELL J J, ZHANG L, ZHOU H, et al. Variations of oral microbiota are associated with pancreatic diseases including pancreatic cancer[J]. Gut, 2012, 61(4): 582-588.
KSIAZEK M, MIZGALSKA D, ENGHILD J J, et al. Miropin, a novel bacterial serpin from the periodontopathogen Tannerella forsythia, inhibits a broad range of proteases by using different peptide bonds within the reactive center loop[J]. J Biol Chem, 2015, 290(1): 658-670.
IKEZOE T. Advances in the diagnosis and treatment of disseminated intravascular coagulation in haematological malignancies[J]. Int J Hematol, 2021, 113(1): 34-44.
YANG L, MANITHODY C, QURESHI S H, et al. Contribution of exosite occupancy by heparin to the regulation of coagulation proteases by antithrombin[J]. Thromb Haemost, 2010, 103(2): 277-283.
SCHECHTER I, BERGER A. On the active site of proteases. 3. Mapping the active site of papain; specific peptide inhibitors of papain[J]. Biochem Biophys Res Commun, 1968, 32(5): 898-902.
SPENCE M A, MORTIMER M D, BUCKLE A M, et al. A comprehensive phylogenetic analysis of the serpin superfamily[J]. Mol Biol Evol, 2021, 38(7): 2915-2929.
Collaborative Computational Project, Number 4. The CCP4 suite: programs for protein crystallography[J]. Acta Crystallogr D Biol Crystallogr, 1994, 50(pt 5): 760-763.
MURSHUDOV G N, SKUBÁK P, LEBEDEV A A, et al. REFMAC5 for the refinement of macromolecular crystal structures[J]. Acta Crystallogr D Biol Crystallogr, 2011, 67(pt 4): 355-367.
MAGGIORA L L, SMITH C W, ZHANG Z Y. A general method for the preparation of internally quenched fluorogenic protease substrates using solid-phase peptide synthesis[J]. J Med Chem, 1992, 35(21): 3727-3730.
WEI Z, YAN Y, CARRELL R W, et al. Crystal structure of protein Z-dependent inhibitor complex shows how protein Z functions as a cofactor in the membrane inhibition of factor X[J]. Blood, 2009, 114(17): 3662-3667.
IRVING J A, CABRITA L D, ROSSJOHN J, et al. The 1.5 Å crystal structure of a prokaryote serpin: controlling conformational change in a heated environment[J]. Structure, 2003, 11(4): 387-397.
IRVING J A, PIKE R N, DAI W, et al. Evidence that serpin architecture intrinsically supports papain-like cysteine protease inhibition: engineering α1-antitrypsin to inhibit cathepsin proteases[J]. Biochemistry, 2002, 41(15): 4998-5004.
CABRITA L D, IRVING J A, PEARCE M C, et al. Aeropin from the extremophile Pyrobaculum aerophilum bypasses the serpin misfolding trap[J]. J Biol Chem, 2007, 282(37): 26802-26809.
TANAKA S, KOGA Y, TAKANO K, et al. Inhibition of chymotrypsin- and subtilisin-like serine proteases with Tk-serpin from hyperthermophilic archaeon Thermococcuskodakaraensis[J]. Biochim Biophys Acta, 2011, 1814(2): 299-307.
LAWRENCE D A, OLSON S T, MUHAMMAD S, et al. Partitioning of serpin-proteinase reactions between stable inhibition and substrate cleavage is regulated by the rate of serpin reactive center loop insertion into β-sheet A[J]. J Biol Chem, 2000, 275(8): 5839-5844.
OWEN M C, BRENNAN S O, LEWIS J H, et al. Mutation of antitrypsin to antithrombin: α1-antitrypsin Pittsburgh (358 Met leads to Arg), a fatal bleeding disorder[J]. N Engl J Med, 1983, 309(12): 694-698.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}