Phosphatidylethanolamine promotes macrophage senescence and liver injury by activating endoplasmic reticulum stress
HAN Longchuan1,2, LI Yue1,2, ZOU Zhihui1,2, LUO Jing1,2,3, LI Ruoyi1, ZHANG Yingting1,2,4, TANG Xinxin1,2, TIAN Lihong1,2,3, LU Yuheng1,2, HUANG Ying1, HE Ming,1,2, FU Yinkun,1,2
1.Frontier Research Center for Cell Homeostasis and Disease Control; Key Laboratory of Cell Differentiation and Apoptosis of Chinese Ministry of Education; Department of Pathophysiology, Shanghai Jiao Tong University College of Basic Medical Sciences, Shanghai 200025, China
2.Institute for Translational Medicine on Cell Fate and Disease, Shanghai Ninth People's Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai 200025, China
3.Department of Pathology and Pathophysiology, School of Basic Medicine, Kunming Medical University, Kunming 650500, China
4.Central Laboratory, Ruijin Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai 200025, China
目的·探讨磷脂酰乙醇胺(phosphatidylethanolamine,PE)对巨噬细胞衰老及其衰老相关分泌表型的影响和分子机制,以及PE在肝损伤中的病理生理学意义。方法·利用阿霉素建立巨噬细胞衰老模型,并给予PE处理。通过腹腔联合注射PE和脂多糖构建小鼠肝损伤模型,观察PE对肝损伤的影响。采用衰老相关β-半乳糖苷酶(senescence-associated β- galactosidase,SA-β-gal)染色,结合实时荧光定量PCR、Western blotting等检测细胞周期抑制蛋白p21、肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)和白介素-6(interleukin-6,IL-6)等衰老标志物及衰老相关分泌表型生物活性因子的表达水平。通过RNA测序结合基因本体论(Gene Ontology,GO)细胞组分富集分析、京都基因与基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)通路富集分析、基因集变异分析(Gene Set Variation Analysis,GSVA)和基因集富集分析(Gene Set Enrichment Analysis,GSEA)筛选PE促进巨噬细胞衰老的信号通路及分子机制。通过体内和体外实验检测内质网应激相关通路中肌醇需求酶1α(inositol requiring enzyme 1 α,IRE1α)、剪接型X盒结合蛋白1(spliced X box binding protein 1,XBP1s)、转录激活因子6(activating transcription factor 6,ATF6)、ATF4、C/EBP同源蛋白(C/EBP homologous protein,CHOP)的表达。结果·PE显著促进巨噬细胞衰老标志物SA-β-gal、p21、p16及衰老相关分泌表型生物活性因子的表达。RNA测序分析显示内质网应激参与PE促进衰老相关分泌表型表达的作用。进一步的实验表明,PE通过激活巨噬细胞内质网应激信号通路促进巨噬细胞衰老及衰老相关分泌表型表达。体内实验证实PE通过内质网应激加剧脂多糖诱导的小鼠肝损伤。结论·PE通过激活内质网应激信号通路,促进巨噬细胞衰老及衰老相关分泌表型生物活性因子分泌,进而加重脂多糖诱导的肝损伤。
关键词:磷脂酰乙醇胺
;
巨噬细胞
;
衰老相关分泌表型
;
肝损伤
;
内质网应激
Abstract
Objective ·To investigate the effects and molecular mechanisms of phosphatidylethanolamine (PE) on macrophage senescence and its senescence-associated secretory phenotype (SASP), as well as its pathophysiological role in liver injury. Methods ·A macrophage senescence model was established using doxorubicin (DOX), followed by PE treatment. A mouse liver injury model was generated via intraperitoneal co-administration of PE and lipopolysaccharide (LPS) to investigate the effects of PE on liver injury. Senescence markers and SASP factors, including senescence-associated β-galactosidase (SA-β-gal), cell cycle inhibitor p21, tumor necrosis factor-α (TNF-α), and interleukin-6 (IL-6), were evaluated using SA-β-gal staining, quantitative real-time PCR, and Western blotting. RNA sequencing (RNA-seq) was performed, followed by Gene Ontology (GO) cellular component enrichment analysis, Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis, Gene Set Variation Analysis (GSVA), and Gene Set Enrichment Analysis (GSEA), to explore the molecular mechanisms and signaling pathways by which PE promotes macrophage senescence. The expression of endoplasmic reticulum (ER) stress-related proteins, including inositol-requiring enzyme 1 α (IRE1α), spliced X-box binding protein 1 (XBP1s), activating transcription factor 6 (ATF6), ATF4, and C/EBP homologous protein (CHOP), was analyzed through in vivo and in vitro experiments. Results ·PE significantly promoted the expression of senescence markers SA-β-gal, p21, p16 and SASP factors. RNA-seq analysis revealed that ER stress was involved in PE-induced promotion of SASP. Further experiments demonstrated that PE activated the ER stress signaling pathway, promoting macrophage senescence and the expression of SASP factors. In vivo experiments further confirmed that PE exacerbated LPS-induced liver injury in mice through ER stress. Conclusion ·PE promotes macrophage senescence and the expression of SASP factors by activating ER stress signaling pathway, thereby aggravating LPS-induced liver injury.
HAN Longchuan, LI Yue, ZOU Zhihui, LUO Jing, LI Ruoyi, ZHANG Yingting, TANG Xinxin, TIAN Lihong, LU Yuheng, HUANG Ying, HE Ming, FU Yinkun. Phosphatidylethanolamine promotes macrophage senescence and liver injury by activating endoplasmic reticulum stress. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2025, 45(6): 693-704 doi:10.3969/j.issn.1674-8115.2025.06.004
小鼠单核巨噬细胞白血病细胞RAW264.7购自美国模式菌种收集中心(American Type Culture Collection,ATCC)。将细胞培养于含10%胎牛血清的DMEM培养基中。由于RAW264.7细胞容易发生自分化,传代培养过程中应避免机械刺激或胰酶处理,并严格控制细胞代数(5~20代);同时应监测细胞的形态变化,若发生明显分化,及时更换新批次细胞[17-18]。
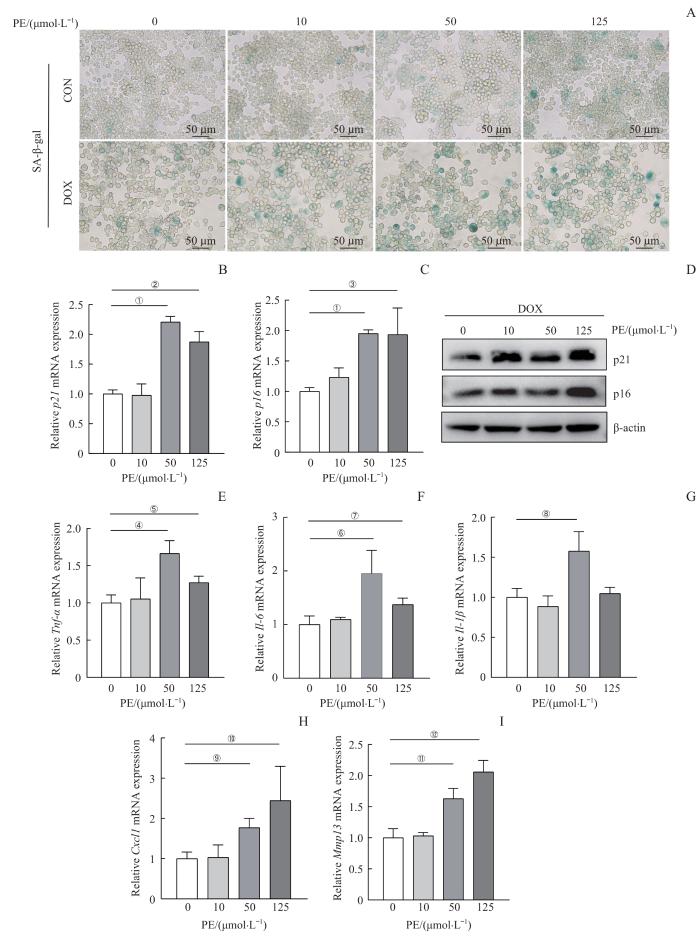
Note: A. Representative images of SA-β-gal staining in RAW264.7 cells treated with different concentrations of PE (0, 10, 50 and 125 μmol·L-1) in the presence or absence of DOX (0.5 μmol·L-1), bar=50 μm. B/C. Quantitative real-time PCR analysis of expression of senescence markers p21 (B) and p16 (C) in RAW264.7 cells treated with different concentrations of PE (0, 10, 50 and 125 μmol·L-1) in the presence of DOX. D. Western blotting analysis of senescence markers. E‒I. Quantitative real-time PCR analysis of expression of senescence-associated secretory phenotype (SASP) factors Tnf-α (E), Il-6 (F), Il-1β (G), Cxcl1 (H) and Mmp13 (I). ①P<0.001, ②P=0.001, ③P=0.021, ④P=0.005, ⑤P=0.027, ⑥P=0.023, ⑦P=0.031, ⑧P=0.019, ⑨P=0.009, ⑩P=0.045, ⑪P=0.008, ⑫P=0.002. DOX—doxorubicin.
Fig 1
PE promotes cellular senescence and the expression of SASP in macrophages
2.2 PE激活内质网应激信号通路
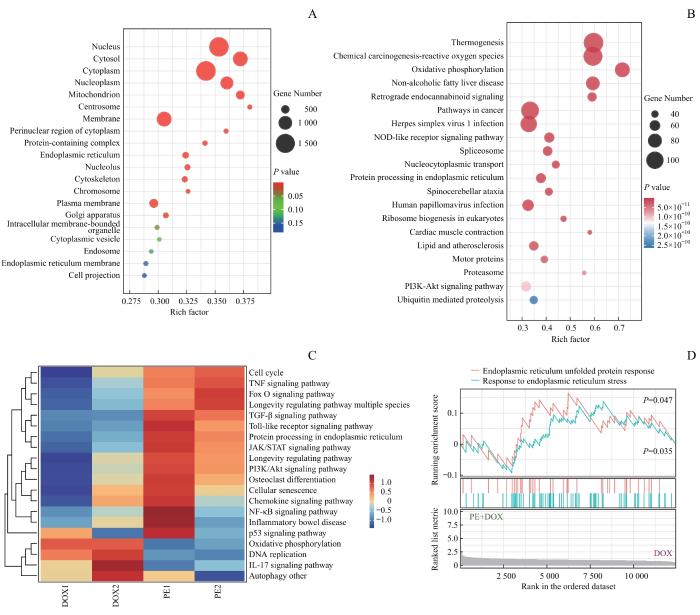
为探究PE促进巨噬细胞衰老的机制,选取50 μmol/L PE联合阿霉素处理的RAW264.7细胞,以及单独使用阿霉素处理的细胞,进行转录组测序。基因本体论(Gene Ontology,GO)细胞组分富集分析显示,差异表达基因的亚细胞定位主要集中在线粒体、内质网和细胞骨架等细胞器(图2A)。京都基因与基因组(Kyoto Encyclopedia of Genes and Genomes,KEGG)通路富集分析进一步表明,经PE处理的衰老巨噬细胞氧化磷酸化和内质网应激相关通路内质网蛋白加工等多条通路显著富集(图2B)。基因集变异分析(Gene Set Variation Analysis,GSVA)热图显示,PE+DOX组在细胞衰老、内质网蛋白加工、PI3K-Akt信号通路、炎症相关通路(如Toll样受体信号通路和NF-κB信号通路)相关基因表达显著增加(图2C)。基因集富集分析(Gene Set Enrichment Analysis,GSEA)显示,PE+DOX组内质网应激相关基因集呈现富集趋势,包括内质网未折叠蛋白反应、内质网应激反应,表明PE处理可能通过增强内质网应激反应驱动巨噬细胞的基因表达变化(图2D)。
Note: The DOX group was treated with 0.5 μmol·L-1 DOX, while the PE group was treated with 50 μmol·L-1 PE in combination with DOX. A. GO enrichment analysis of cellular component categories, presented as an enrichment factor plot [(PE+DOX) vs DOX]. B. KEGG pathway enrichment analysis displayed as an enrichment factor plot [(PE+DOX) vs DOX]. C. Heatmap of GSVA [(PE+DOX) vs DOX]. D. GSEA of endoplasmic reticulum unfolded protein response and response to endoplasmic reticulum stress [(PE+DOX) vs DOX].
Fig 2
Functional enrichment analysis of RNA sequencing data from senescent macrophages treated with PE
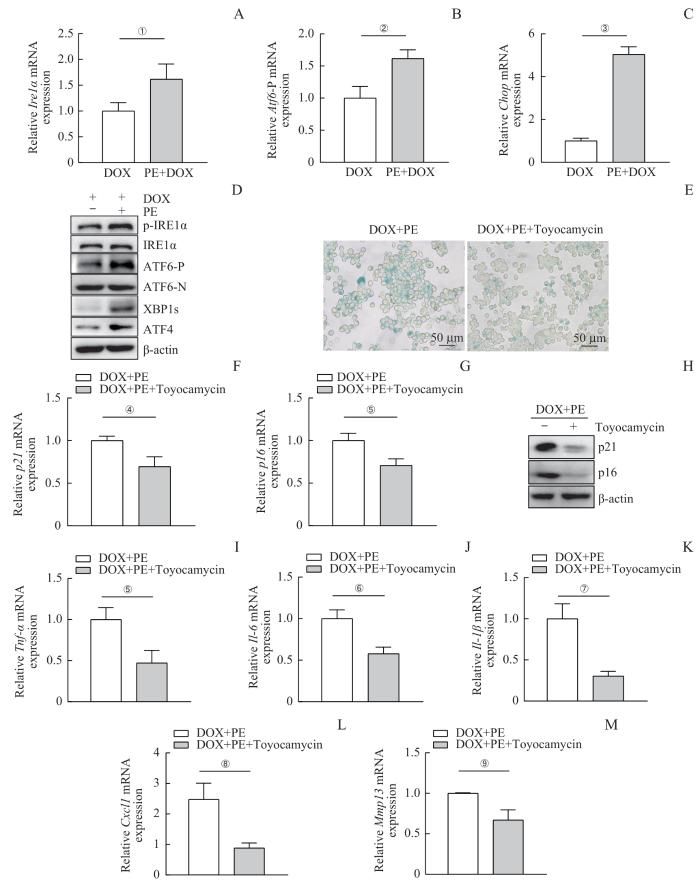
Note:A‒C. Quantitative real-time PCR analysis of Ire1α (A), Atf6 (B) and Chop (C) mRNA expression in RAW264.7 cells treated with DOX and PE (50 μmol·L-1). D. Western blotting analysis of endoplasmic reticulum stress-related proteins, including p-IRE1α, IRE1α, ATF6-P, ATF6-N, XBP1s and ATF4. E. SA-β-gal staining of RAW264.7 cells treated with Toyocamycin, DOX and PE. F/G. Quantitative real-time PCR analysis of senescence marker mRNA expression, including p21 (F) and p16 (G). H. Western blotting analysis of proteins including p21 and p16. I‒M. Quantitative real-time PCR analysis of SASP mRNA expression, including Tnf-α (I), Il-6 (J), Il-1β (K), Cxcl1 (L) and Mmp13 (M). ①P=0.033, ②P=0.009, ③P<0.001, ④P=0.014, ⑤P=0.012, ⑥P=0.005, ⑦P=0.003, ⑧P=0.008, ⑨P=0.010.
Fig 3
PE promotes macrophage senescence and SASP through activating endoplasmic reticulum stress signaling pathway
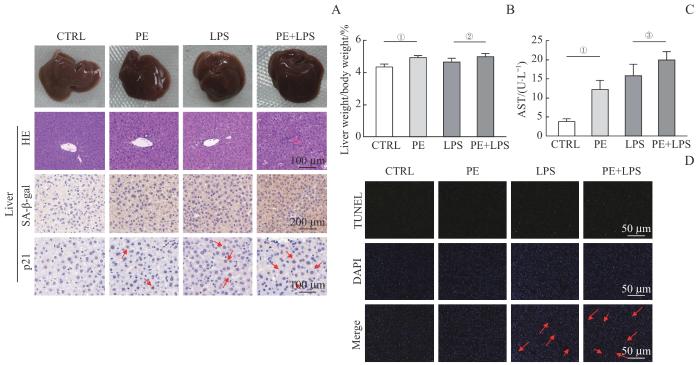
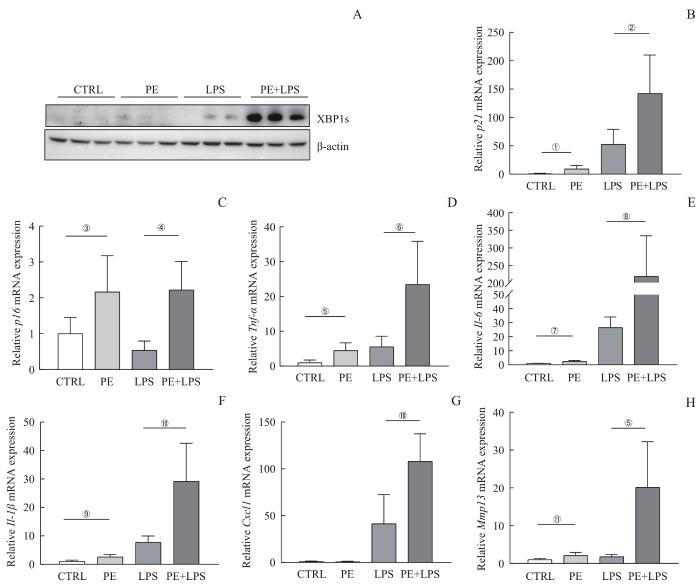
Note: Mice were divided into the PE group, lipopolysaccharide (LPS) group, and PE+LPS group by intraperitoneal injection of PE, LPS, and PE combined with LPS, respectively, with an additional blank control (CTRL) group. A. General morphology, HE staining and immunohistochemistry of the liver from the four groups of mice. HE staining, bar=100 μm; SA-β-gal immunohistochemistry, bar=200 μm; p21 immunohistochemistry, bar=100 μm. Red arrows indicate p21-positive cells. B. Percentages of liver weight. C. Plasma AST in the indicated mice. D. TUNEL staining of liver, bar=50 μm. Red arrows indicate TUNEL-positive cells. ①P<0.001, ②P=0.041, ③P=0.037.
Fig 4
PE exacerbates lipopolysaccharide-induced liver injury
内质网应激被认为是衰老过程中一个重要的分子机制,其通过未折叠蛋白反应(unfolded protein response,UPR)启动一系列细胞应激反应。PE可能通过加强内质网应激促使巨噬细胞向衰老表型转变,进而加剧局部炎症反应。巨噬细胞衰老相关分泌表型的主要生物活性因子之一IL-6是肝细胞增生过程中重要的启动因子,同时也是激活信号转导和转录激活因子3(signal transducer and activator of transcription 3,STAT3)的主要细胞因子[27]。在内质网应激过程中,IRE1α的表达上调能激活细胞中IL-6的表达并会随着IRE1α-XBPl途径中XBP1剪切形式XBP1s的增加而显著增加[28]。转录组测序结果显示,PE显著激活内质网应激相关信号通路。PE通过促进内质网应激,并可能是通过诱导内质网功能失调来改变巨噬细胞的生理状态,从而促进其衰老进程。由于PE处理显著促进RAW264.7细胞发生内质网应激,而且在内质网应激所有的信号通路中,IRE1α-XBP1信号通路变化最为显著,因此本研究利用XBP1s特异性抑制剂丰加霉素证实PE主要通过激活XBP1信号通路促进巨噬细胞衰老及衰老相关分泌表型表达。本研究参考相关的研究[29-30]并结合预实验结果发现,丰加霉素在高浓度(1 μmol/L)的情况下会抑制细胞增殖,随时间延长发生细胞死亡,而低浓度的情况下(1~100 nmol/L)对细胞没有明显毒性。因此,本研究在实验中使用20 nmol/L处理细胞且没有发现细胞死亡,XBP1s也得到有效抑制。这种实验设计既确保了XBP1通路抑制的有效性,又排除了细胞毒性对实验结果的影响。本研究结果表明PE是通过内质网应激通路调节巨噬细胞功能的一个关键分子,为进一步探索PE在细胞衰老和免疫调控中的作用提供了重要的数据支持。
This study was reviewed and approved by Institutional Animal Care and Use Committee (IACUC) of Shanghai Jiao Tong University School of Medicine (Approval Letter No. JUMC2023-173-A). All experimental animal protocols in this study were conducted in accordance with the Guidelines for Animal Welfare and Use of Experimental Animals of Shanghai Jiao Tong University School of Medicine.
HAN Longchuan was primarily responsible for conducting experiments, organizing and analyzing data, and drafting and revising the manuscript. LI Yue contributed to experimental operations and manuscript revision. ZOU Zhihui and LUO Jing participated in data analysis and manuscript revisions. LI Ruoyi contributed to animal experiments and manuscript revisions. ZHANG Yingting, TANG Xinxin, TIAN Lihong, LU Yuheng and HUANG Ying participated in manuscript revisions. HE Ming and FU Yinkun were responsible for project design and manuscript revisions. All authors have read the final version of the paper and agreed to its submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
ZHANG L, PITCHER L E, YOUSEFZADEH M J, et al. Cellular senescence: a key therapeutic target in aging and diseases[J]. J Clin Invest, 2022, 132(15): e158450.
GUO J, HUANG X Q, DOU L, et al. Aging and aging-related diseases: from molecular mechanisms to interventions and treatments[J]. Signal Transduct Target Ther, 2022, 7(1): 391.
PEISELER M, DAVID B A, ZINDEL J, et al. Kupffer cell-like syncytia replenish resident macrophage function in the fibrotic liver[J]. Science, 2023, 381(6662): eabq5202.
ZHAO B H, WU B, FENG N, et al. Aging microenvironment and antitumor immunity for geriatric oncology: the landscape and future implications[J]. J Hematol Oncol, 2023, 16(1): 28.
LEON K E, BUJ R, LESKO E, et al. DOT1L modulates the senescence-associated secretory phenotype through epigenetic regulation of IL1A[J]. J Cell Biol, 2021, 220(8): e202008101.
WANG B S, HAN J, ELISSEEFF J H, et al. The senescence-associated secretory phenotype and its physiological and pathological implications[J]. Nat Rev Mol Cell Biol, 2024, 25(12): 958-978.
YOSHIMOTO S, LOO T M, ATARASHI K, et al. Obesity-induced gut microbial metabolite promotes liver cancer through senescence secretome[J]. Nature, 2013, 499(7456): 97-101.
LIU X, HARTMAN C L, LI L Y, et al. Reprogramming lipid metabolism prevents effector T cell senescence and enhances tumor immunotherapy[J]. Sci Transl Med, 2021, 13(587): eaaz6314.
GIBELLINI F, SMITH T K. The Kennedy pathway: de novo synthesis of phosphatidylethanolamine and phosphatidylcholine[J]. IUBMB Life, 2010, 62(6): 414-428.
TACIAK B, BIAŁASEK M, BRANIEWSKA A, et al. Evaluation of phenotypic and functional stability of RAW 264.7 cell line through serial passages[J]. PLoS One, 2018, 13(6): e0198943.
FACCHIN B M, DOS REIS G O, VIEIRA G N, et al. Inflammatory biomarkers on an LPS-induced RAW 264.7 cell model: a systematic review and meta-analysis[J]. Inflamm Res, 2022, 71(7/8): 741-758.
YEN L, SVENDSEN J, LEE J S, et al. Exogenous control of mammalian gene expression through modulation of RNA self-cleavage[J]. Nature, 2004, 431(7007): 471-476.
EL MANAA W, DUPLAN E, GOIRAN T, et al. Transcription- and phosphorylation-dependent control of a functional interplay between XBP1s and PINK1 governs mitophagy and potentially impacts Parkinson disease pathophysiology[J]. Autophagy, 2021, 17(12): 4363-4385.
ZHANG G Z, ZHAN M S, ZHANG C C, et al. Redox-responsive dendrimer nanogels enable ultrasound-enhanced chemoimmunotherapy of pancreatic cancer via endoplasmic reticulum stress amplification and macrophage polarization[J]. Adv Sci (Weinh), 2023, 10(24): e2301759.
XUE L J, LIU K, YAN C X, et al. Schisandra lignans ameliorate nonalcoholic steatohepatitis by regulating aberrant metabolism of phosphatidylethanolamines[J]. Acta Pharm Sin B, 2023, 13(8): 3545-3560.
ZHANG L Z, RICHARD A S, JACKSON C B, et al. Phosphatidylethanolamine and phosphatidylserine synergize to enhance GAS6/AXL-mediated virus infection and efferocytosis[J]. J Virol, 2020, 95(2): e02079-20.
VAZQUEZ-DE-LARA L G, TLATELPA-ROMERO B, ROMERO Y, et al. Phosphatidylethanolamine induces an antifibrotic phenotype in normal human lung fibroblasts and ameliorates bleomycin-induced lung fibrosis in mice[J]. Int J Mol Sci, 2018, 19(9): 2758.
HUANG F Z, LIU X M, LIU J J, et al. Phosphatidylethanolamine aggravates Angiotensin Ⅱ-induced atrial fibrosis by triggering ferroptosis in mice[J]. Front Pharmacol, 2023, 14: 1148410.
LI L, CUI L, LIN P, et al. Kupffer-cell-derived IL-6 is repurposed for hepatocyte dedifferentiation via activating progenitor genes from injury-specific enhancers[J]. Cell Stem Cell, 2023, 30(3): 283-299.e9.
FANG P P, XIANG L X, HUANG S S, et al. IRE1α-XBP1 signaling pathway regulates IL-6 expression and promotes progression of hepatocellular carcinoma[J]. Oncol Lett, 2018, 16(4): 4729-4736.
RI M, TASHIRO E, OIKAWA D, et al. Identification of Toyocamycin, an agent cytotoxic for multiple myeloma cells, as a potent inhibitor of ER stress-induced XBP1 mRNA splicing[J]. Blood Cancer J, 2012, 2(7): e79.
SONG N, SONG Y Q, HU B B, et al. Persistent endoplasmic reticulum stress stimulated by peptide assemblies for sensitizing cancer chemotherapy[J]. Adv Healthcare Mater, 2023, 12(5): 2202039.
YIN N, ZHANG W J, SUN X X, et al. Artificial cells delivering itaconic acid induce anti-inflammatory memory-like macrophages to reverse acute liver failure and prevent reinjury[J]. Cell Rep Med, 2023, 4(8): 101132.
... 内质网应激被认为是衰老过程中一个重要的分子机制,其通过未折叠蛋白反应(unfolded protein response,UPR)启动一系列细胞应激反应.PE可能通过加强内质网应激促使巨噬细胞向衰老表型转变,进而加剧局部炎症反应.巨噬细胞衰老相关分泌表型的主要生物活性因子之一IL-6是肝细胞增生过程中重要的启动因子,同时也是激活信号转导和转录激活因子3(signal transducer and activator of transcription 3,STAT3)的主要细胞因子[27].在内质网应激过程中,IRE1α的表达上调能激活细胞中IL-6的表达并会随着IRE1α-XBPl途径中XBP1剪切形式XBP1s的增加而显著增加[28].转录组测序结果显示,PE显著激活内质网应激相关信号通路.PE通过促进内质网应激,并可能是通过诱导内质网功能失调来改变巨噬细胞的生理状态,从而促进其衰老进程.由于PE处理显著促进RAW264.7细胞发生内质网应激,而且在内质网应激所有的信号通路中,IRE1α-XBP1信号通路变化最为显著,因此本研究利用XBP1s特异性抑制剂丰加霉素证实PE主要通过激活XBP1信号通路促进巨噬细胞衰老及衰老相关分泌表型表达.本研究参考相关的研究[29-30]并结合预实验结果发现,丰加霉素在高浓度(1 μmol/L)的情况下会抑制细胞增殖,随时间延长发生细胞死亡,而低浓度的情况下(1~100 nmol/L)对细胞没有明显毒性.因此,本研究在实验中使用20 nmol/L处理细胞且没有发现细胞死亡,XBP1s也得到有效抑制.这种实验设计既确保了XBP1通路抑制的有效性,又排除了细胞毒性对实验结果的影响.本研究结果表明PE是通过内质网应激通路调节巨噬细胞功能的一个关键分子,为进一步探索PE在细胞衰老和免疫调控中的作用提供了重要的数据支持. ...
1
... 内质网应激被认为是衰老过程中一个重要的分子机制,其通过未折叠蛋白反应(unfolded protein response,UPR)启动一系列细胞应激反应.PE可能通过加强内质网应激促使巨噬细胞向衰老表型转变,进而加剧局部炎症反应.巨噬细胞衰老相关分泌表型的主要生物活性因子之一IL-6是肝细胞增生过程中重要的启动因子,同时也是激活信号转导和转录激活因子3(signal transducer and activator of transcription 3,STAT3)的主要细胞因子[27].在内质网应激过程中,IRE1α的表达上调能激活细胞中IL-6的表达并会随着IRE1α-XBPl途径中XBP1剪切形式XBP1s的增加而显著增加[28].转录组测序结果显示,PE显著激活内质网应激相关信号通路.PE通过促进内质网应激,并可能是通过诱导内质网功能失调来改变巨噬细胞的生理状态,从而促进其衰老进程.由于PE处理显著促进RAW264.7细胞发生内质网应激,而且在内质网应激所有的信号通路中,IRE1α-XBP1信号通路变化最为显著,因此本研究利用XBP1s特异性抑制剂丰加霉素证实PE主要通过激活XBP1信号通路促进巨噬细胞衰老及衰老相关分泌表型表达.本研究参考相关的研究[29-30]并结合预实验结果发现,丰加霉素在高浓度(1 μmol/L)的情况下会抑制细胞增殖,随时间延长发生细胞死亡,而低浓度的情况下(1~100 nmol/L)对细胞没有明显毒性.因此,本研究在实验中使用20 nmol/L处理细胞且没有发现细胞死亡,XBP1s也得到有效抑制.这种实验设计既确保了XBP1通路抑制的有效性,又排除了细胞毒性对实验结果的影响.本研究结果表明PE是通过内质网应激通路调节巨噬细胞功能的一个关键分子,为进一步探索PE在细胞衰老和免疫调控中的作用提供了重要的数据支持. ...
1
... 内质网应激被认为是衰老过程中一个重要的分子机制,其通过未折叠蛋白反应(unfolded protein response,UPR)启动一系列细胞应激反应.PE可能通过加强内质网应激促使巨噬细胞向衰老表型转变,进而加剧局部炎症反应.巨噬细胞衰老相关分泌表型的主要生物活性因子之一IL-6是肝细胞增生过程中重要的启动因子,同时也是激活信号转导和转录激活因子3(signal transducer and activator of transcription 3,STAT3)的主要细胞因子[27].在内质网应激过程中,IRE1α的表达上调能激活细胞中IL-6的表达并会随着IRE1α-XBPl途径中XBP1剪切形式XBP1s的增加而显著增加[28].转录组测序结果显示,PE显著激活内质网应激相关信号通路.PE通过促进内质网应激,并可能是通过诱导内质网功能失调来改变巨噬细胞的生理状态,从而促进其衰老进程.由于PE处理显著促进RAW264.7细胞发生内质网应激,而且在内质网应激所有的信号通路中,IRE1α-XBP1信号通路变化最为显著,因此本研究利用XBP1s特异性抑制剂丰加霉素证实PE主要通过激活XBP1信号通路促进巨噬细胞衰老及衰老相关分泌表型表达.本研究参考相关的研究[29-30]并结合预实验结果发现,丰加霉素在高浓度(1 μmol/L)的情况下会抑制细胞增殖,随时间延长发生细胞死亡,而低浓度的情况下(1~100 nmol/L)对细胞没有明显毒性.因此,本研究在实验中使用20 nmol/L处理细胞且没有发现细胞死亡,XBP1s也得到有效抑制.这种实验设计既确保了XBP1通路抑制的有效性,又排除了细胞毒性对实验结果的影响.本研究结果表明PE是通过内质网应激通路调节巨噬细胞功能的一个关键分子,为进一步探索PE在细胞衰老和免疫调控中的作用提供了重要的数据支持. ...
1
... 内质网应激被认为是衰老过程中一个重要的分子机制,其通过未折叠蛋白反应(unfolded protein response,UPR)启动一系列细胞应激反应.PE可能通过加强内质网应激促使巨噬细胞向衰老表型转变,进而加剧局部炎症反应.巨噬细胞衰老相关分泌表型的主要生物活性因子之一IL-6是肝细胞增生过程中重要的启动因子,同时也是激活信号转导和转录激活因子3(signal transducer and activator of transcription 3,STAT3)的主要细胞因子[27].在内质网应激过程中,IRE1α的表达上调能激活细胞中IL-6的表达并会随着IRE1α-XBPl途径中XBP1剪切形式XBP1s的增加而显著增加[28].转录组测序结果显示,PE显著激活内质网应激相关信号通路.PE通过促进内质网应激,并可能是通过诱导内质网功能失调来改变巨噬细胞的生理状态,从而促进其衰老进程.由于PE处理显著促进RAW264.7细胞发生内质网应激,而且在内质网应激所有的信号通路中,IRE1α-XBP1信号通路变化最为显著,因此本研究利用XBP1s特异性抑制剂丰加霉素证实PE主要通过激活XBP1信号通路促进巨噬细胞衰老及衰老相关分泌表型表达.本研究参考相关的研究[29-30]并结合预实验结果发现,丰加霉素在高浓度(1 μmol/L)的情况下会抑制细胞增殖,随时间延长发生细胞死亡,而低浓度的情况下(1~100 nmol/L)对细胞没有明显毒性.因此,本研究在实验中使用20 nmol/L处理细胞且没有发现细胞死亡,XBP1s也得到有效抑制.这种实验设计既确保了XBP1通路抑制的有效性,又排除了细胞毒性对实验结果的影响.本研究结果表明PE是通过内质网应激通路调节巨噬细胞功能的一个关键分子,为进一步探索PE在细胞衰老和免疫调控中的作用提供了重要的数据支持. ...



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}