SIRT2 regulates macrophage chemotaxis by de-modifying histone H4K8 lactylation
SONG Wenting,1, TAO Yue2, PAN Yi2, MO Xi2, CAO Qing,1
1.Department of Infectious Disease, Shanghai Children´s Medical Center, Shanghai Jiao Tong University School of Medicine, Shanghai 200127, China
2.Pediatric Translational Medicine Institute, Shanghai Children´s Medical Center, Shanghai Jiao Tong University School of Medicine, Shanghai 200127, China
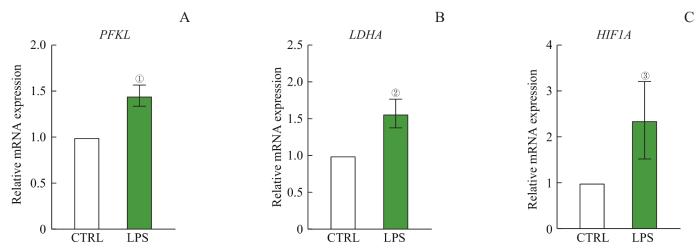
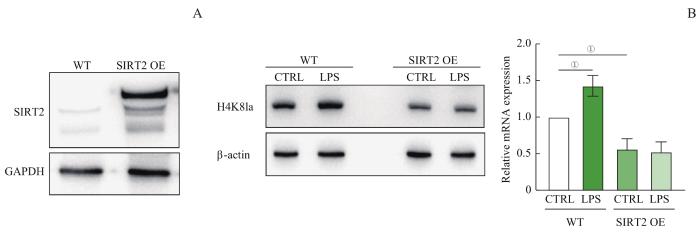
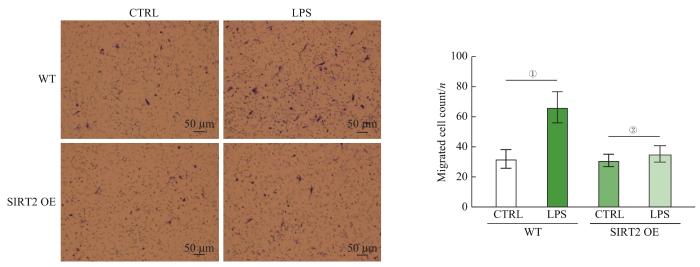
Objective ·To explore the regulatory role of silent information regulator 2 (SIRT2) in modulating the immune phenotype of macrophages after infection by removing the lactylation at H4K8 site of histone and the corresponding mechanism. Methods ·Human THP-1 leukemia cells were induced by phorbol 12-myristate 13-acetate (PMA) and stimulated by lipopolysaccharide (LPS) to establish a macrophage infection model. Macrophages without LPS treatment (pTHP-1) were set as the control (CTRL) group, and macrophages with LPS treatment were set as the infected (LPS) group. Western blotting was used to detect the level of histone modification and SIRT2 protein in macrophages. RT-qPCR was used to detect the expression level of glycolytic key enzymes [phosphofructokinase liver type (PFKL), lactate dehydrogenase A (LDHA)] and modulators genes hypoxia inducible factor 1α (HIF-1α), and the expression level of Sirtuin genes and HDAC genes between the two groups. Transwell was used to detect the ability of macrophage chemotaxis. Lentivirus packaging and cell infection were used to construct SIRT2 overexpression cell line. The interaction analysis method of RNA sequencing (RNA-seq) and chromatin immunoprecipitation sequencing (ChIP-seq) was used to analyze the difference and pathway enrichment of the genes specifically bound to H4K8 lactylation (H4K8la). Results ·Compared to the CTRL group, macrophage glycolysis was upregulated and the level of H4K8la was significantly increased in the LPS group (P<0.05), while the level of lactylation in other sites remained unchanged. Among all known enzymes with deacetylation modification function, only SIRT2 showed a significant decrease after LPS treatment (P<0.05), and overexpression of SIRT2 could significantly inhibit the level of H4K8la modification, while the level of H4K8ac remained unchanged (P>0.05). The interactive analysis of ChIP-seq and RNA-seq revealed that chemotaxis-related genes were regulated by H4K8la, and macrophage chemotaxis ability significantly decreased after the overexpression of SIRT2 and downregulation of H4K8la (P<0.05). Conclusion ·SIRT2 can change the expression of target genes related to chemotaxis by removing H4K8la modification, thereby reducing the chemotaxis ability of macrophages. Targeting SIRT2 and H4K8la modification may help control inflammation mediated by macrophages.
SONG Wenting, TAO Yue, PAN Yi, MO Xi, CAO Qing. SIRT2 regulates macrophage chemotaxis by de-modifying histone H4K8 lactylation. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(8): 1008-1016 doi:10.3969/j.issn.1674-8115.2023.08.008
RNA测序(RNA sequencing,RNA-seq)文库构建和Illumina测序由明码生物科技(上海)有限公司完成;使用基因本体(gene ontology,GO)数据库、京都基因和基因组百科全书(Kyoto Encyclopedia of Genes and Genomes,KEGG)数据库进行基因功能注释;使用DESeq2软件分析各细胞之间的差异基因表达。
1.3.7 ChIP测序
ChIP测序技术(chromatin immunoprecipitation sequencing,ChIP-seq)是ChIP后结合高通量测序的方法。取LPS组巨噬细胞,使用1%甲醛交联10 min后加入1×甘氨酸终止交联;SDS裂解液重悬细胞后超声法破碎DNA,取10%DNA产物作为Input,其余DNA产物用于后续免疫沉淀(immunoprecipitation,IP)实验,分别加入抗组蛋白H4第8位赖氨酸乳酸化(lactylation of histone H4 lysine 8,H4K8la)抗体及抗IgG抗体,4 ℃低速混合孵育过夜;每个样本中加入30 μL protein A/G beads,4 ℃低速混合孵育2 h;按说明书洗涤DNA产物,并使用Elute buffer洗脱;加入无水乙醇沉淀产物后,使用乙二胺四乙酸(ethylenediaminetetra-acetic acid,EDTA)、三羟甲基氨基甲烷(trihydroxymethyl aminomethane,Tris)及Proteinase K解交联;使用DNA纯化试剂盒纯化DNA。后续文库构建和测序由安诺优达基因科技有限公司完成。
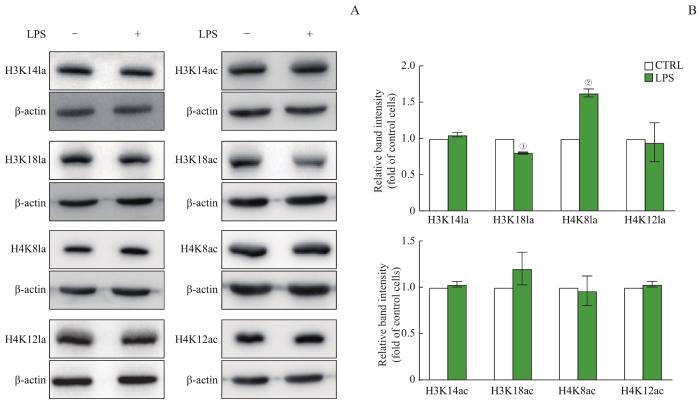
Note: A. Detection of histone lactylation (left) and acetylation (right) levels by Western blotting. B. Expression of histone lactylation (above) and acetylation (below). ①P=0.000,②P=0.002, compared with the CTRL group.
Fig 2
Histone modification levels in macrophages detected by Western blotting
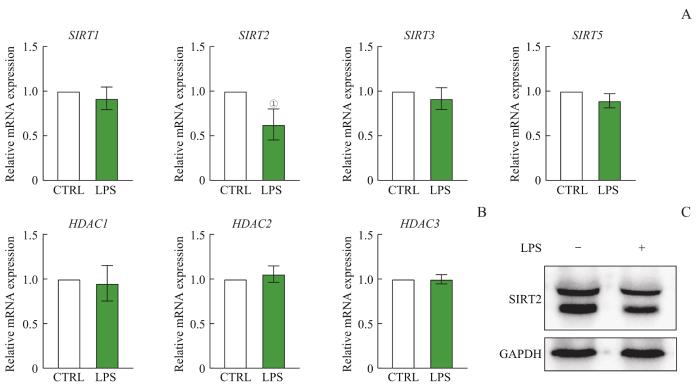
Note: A. Expression of Sirtuin family mRNA. B. Expression of HDAC family mRNA. C. Detection of SIRT2 protein by Western blotting. ①P=0.010, compared with the CTRL group.
Fig 3
RNA expression levels of Sirtuin family and HDAC family of macrophages after LPS treatment
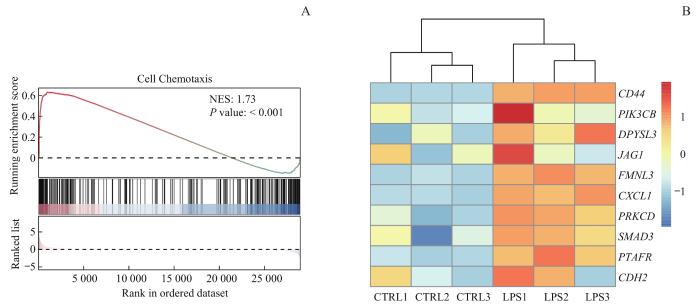
Note: A. Gene set enrichment analysis of macrophages after LPS stimulation. NES—normalized enrichment score. B. Heatmap for macrophage chemotaxis- related genes bound to H4K8la. CTRL1, CTRL2, and CTRL3 were three replicate samples from the control group, while LPS1, LPS2, and LPS3 were three replicate samples from the LPS-treated infection group. PIK3CB—phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit beta; DPYSL3—dihydropyrimidinase like 3; JAG1—jagged canonical Notch ligand 1; FMNL3—formin like 3; CXCL1—C-X-C motif chemokine ligand 1; PRKCD—protein kinase C delta; SMAD3—SMAD family member 3; PTAFR—platelet activating factor receptor; CDH2—cadherin 2.
Fig 6
Interaction analysis of RNA-seq and ChIP-seq
The study was designed by SONG Wenting and MO Xi. The experimental operation was completed by SONG Wenting and PAN Yi. The manuscript was drafted and revised by SONG Wenting, MO Xi, TAO Yue and CAO Qing. All the authors have read the last version of paper and consented for submission.
利益冲突声明
本研究不存在利益冲突。
COMPETING INTERESTS
All authors disclose no relevant conflict of interests.
PETER K, REHLI M, SINGER K, et al. Lactic acid delays the inflammatory response of human monocytes[J]. Biochem Biophys Res Commun, 2015, 457(3): 412-418.
ERREA A, CAYET D, MARCHETTI P, et al. Lactate inhibits the pro-inflammatory response and metabolic reprogramming in murine macrophages in a GPR81-independent manner[J]. PLoS One, 2016, 11(11): e0163694.
NAREIKA A, HE L, GAME B A, et al. Sodium lactate increases LPS-stimulated MMP and cytokine expression in U937 histiocytes by enhancing AP-1 and NF-κB transcriptional activities[J]. Am J Physiol Endocrinol Metab, 2005, 289(4): E534-E542.
SAMUVEL D J, SUNDARARAJ K P, NAREIKA A, et al. Lactate boosts TLR4 signaling and NF-κB pathway-mediated gene transcription in macrophages via monocarboxylate transporters and MD-2 up-regulation[J]. J Immunol, 2009, 182(4): 2476-2484.
XU H W, WU M Y, MA X M, et al. Function and mechanism of novel histone posttranslational modifications in health and disease[J]. Biomed Res Int, 2021, 2021: 6635225.
MOHAMMADI A, SHARIFI A, POURPAKNIA R, et al. Manipulating macrophage polarization and function using classical HDAC inhibitors: implications for autoimmunity and inflammation[J]. Crit Rev Oncol, 2018, 128: 1-18.
NEWSHOLME P, GORDON S, NEWSHOLME E A. Rates of utilization and fates of glucose, glutamine, pyruvate, fatty acids and ketone bodies by mouse macrophages[J]. Biochem J, 1987, 242(3): 631-636.
PARK D, LIM G, YOON S J, et al. The role of immunomodulatory metabolites in shaping the inflammatory response of macrophages[J]. BMB Rep, 2022, 55(11): 519-527.
COLEGIO O R, CHU N Q, SZABO A L, et al. Functional polarization of tumour-associated macrophages by tumour-derived lactic acid[J]. Nature, 2014, 513(7519): 559-563.
DIETL K, RENNER K, DETTMER K, et al. Lactic acid and acidification inhibit TNF secretion and glycolysis of human monocytes[J]. J Immunol, 2010, 184(3): 1200-1209.
ALARCÓN P, MANOSALVA C, CONEJEROS I, et al. D (-) lactic acid-induced adhesion of bovine neutrophils onto endothelial cells is dependent on neutrophils extracellular traps formation and CD11b expression[J]. Front Immunol, 2017, 8: 975.
WU D, SHI Y X, ZHANG H, et al. Epigenetic mechanisms of Immune remodeling in sepsis: targeting histone modification[J]. Cell Death Dis, 2023, 14(2): 112.
CHEN L H, HUANG L X, GU Y, et al. Lactate-lactylation hands between metabolic reprogramming and immunosuppression[J]. Int J Mol Sci, 2022, 23(19): 11943.
IRIZARRY-CARO R A, MCDANIEL M M, OVERCAST G R, et al. TLR signaling adapter BCAP regulates inflammatory to reparatory macrophage transition by promoting histone lactylation[J]. Proc Natl Acad Sci USA, 2020, 117(48): 30628-30638.
KOVACS L, CAO Y P, HAN W H, et al. PFKFB3 in smooth muscle promotes vascular remodeling in pulmonary arterial hypertension[J]. Am J Respir Crit Care Med, 2019, 200(5): 617-627.
WANG N X, WANG W W, WANG X Q, et al. Histone lactylation boosts reparative gene activation post-myocardial infarction[J]. Circ Res, 2022, 131(11): 893-908.
MA W, AO S, ZHOU J, et al. Methylsulfonylmethane protects against lethal dose MRSA-induced sepsis through promoting M2 macrophage polarization[J]. Mol Immunol, 2022, 146: 69-77.
CHU X, DI C Y, CHANG P P, et al. Lactylated histone H3K18 as a potential biomarker for the diagnosis and predicting the severity of septic shock[J]. Front Immunol, 2022, 12: 786666.
DAI H, SINCLAIR D A, ELLIS J L, et al. Sirtuin activators and inhibitors: promises, achievements, and challenges[J]. Pharmacol Ther, 2018, 188: 140-154.
XU H, YU X, WANG B, et al. The clinical significance of the SIRT2 expression level in the early stage of sepsis patients[J]. Ann Palliat Med, 2020, 9(4): 1413-1419.
SASSO G L, MENZIES K J, MOTTIS A, et al. SIRT2 deficiency modulates macrophage polarization and susceptibility to experimental colitis[J]. PLoS One, 2014, 9(7): e103573.
ZU H X, LI C, DAI C R, et al. SIRT2 functions as a histone delactylase and inhibits the proliferation and migration of neuroblastoma cells[J]. Cell Discov, 2022, 8(1): 54.
TU Q Q, YU X Y, XIE W, et al. Prokineticin 2 promotes macrophages-mediated antibacterial host defense against bacterial pneumonia[J]. Int J Infect Dis, 2022, 125: 103-113.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}