Mechanism of DUX-induced differentiation of mESC into extraembryonic endoderm
HONG Lei,1,∗, GUO Chuanliang,1,∗, CAI Qin1, LI Wanrui1, ZENG Yitao1, XUE Yan,1,2, ZENG Fanyi,1,2
1.Shanghai Institute of Medical Genetics, Shanghai Children′s Hospital, Shanghai Jiao Tong University School of Medicine, NHC Key Laboratory of Medical Embryogenesis and Developmental Molecular Biology & Shanghai Key Laboratory of Embryo and Reproduction Engineering, Shanghai 200040, China
2.Department of Histoembryology, Genetics & Development, Shanghai Jiao Tong University College of Basic Medical Sciences, Shanghai 200025, China
目的·探索双同源盒(double homeobox,DUX)蛋白对小鼠胚胎干细胞(mouse embryonic stem cell,mESC)向胚外内胚层(extraembryonic endoderm,XEN)分化潜能的影响及可能的作用机制。方法·使用慢病毒体系在mESC中构建过表达DUX细胞系,利用流式细胞术检测DUX过表达前后2细胞样细胞(2-cell-like cell,2CLC)的比例,并使用实时定量聚合酶链反应(real-time quantitative reverse transcription polymerase chain reaction,RT-qPCR)检测2细胞期特异性基因,如内源性Dux、锌指和SCAN结构域的蛋白质4c(zinc finger and SCAN domain containing 4c,Zscan4c)、锌指蛋白352(zinc finger protein 352,Zfp352)和鼠内源性反转录病毒-聚合酶(murine endogenous retrovirus-L polymerase,MERVL-pol)的表达。RT-qPCR检测过表达DUX的mESC多能性因子[nanog homeobox(Nanog)、kruppel-like transcription factor 4(Klf4)、性别决定区Y框蛋白2(sex determining region Y-box 2,Sox2)、八聚体结合转录因子4(octamer-binding transcription factor 4,Oct4)]和自然分化状态下各胚层标志性基因[内胚层(endodermal):GATA结合蛋白4(GATA binding protein 4,Gata4)、Gata6、Sox17;外胚层(ectodermal):微Ⅲ型β微管蛋白3(tubulin beta 3 class Ⅲ,Tubb3)、巢蛋白(Nestin);中胚层(mesodermal):心脏和神经嵴衍生物表达转录本1(heart and neural crest derivatives expressed 1,Hand1)、肌源性分化蛋白1(myogenic differentiation 1,Myod1)、激酶插入结构域受体(kinase insert domain protein receptor,Flk1)]的表达。挖掘公共转录组测序(RNA sequencing,RNA-seq)数据,通过分析胚外内胚层标志基因的表达水平,明确DUX对mESC向胚外内胚层分化的影响;通过对差异基因的功能及通路进行基因本体论(Gene Ontology,GO)富集分析、京都基因和基因组数据库(Kyoto Encyclopedia of Genes and Genomes,KEGG)富集分析和基因集富集分析(gene set enrichment analysis,GSEA),找出DUX作用的信号通路;深入分析已有的染色质免疫共沉淀技术结合二代测序(chromatin immunoprecipitation sequencing,ChIP-seq)数据,探究DUX的潜在靶基因。结果·2CLC比例升高和2细胞期标志基因表达上调,证明已成功构建过表达DUX细胞系。分子生物学实验显示过表达DUX后可有效维持mESC的多能性,与公共RNA-seq数据分析结果一致;差异基因分析发现,内胚层基因出现特异性上调;诱导mESC自然分化后,RT-qPCR检测实验表明XEN标志基因(Gata4、Gata6、Sox17)的mRNA表达出现显著上调(P<0.001),而中胚层、外胚层基因没有特异性变化。GSEA结果提示DUX可能激活了视黄醇代谢信号通路,ChIP-seq数据解析进一步揭示在DUX结合的peaks中存在已知的视黄酸受体motif,可激活下游与XEN发育相关的靶基因。结论·DUX与视黄酸信号通路密切关联,预示其激活了视黄酸信号通路,促进mESC倾向XEN分化。
关键词:小鼠胚胎干细胞
;
胚外内胚层细胞
;
双同源盒
;
视黄酸信号通路
Abstract
Objective ·To explore the effect of double homeobox (DUX) protein on the differentiation potential of mouse embryonic stem cells (mESCs) into extraembryonic endoderm (XEN) and the possible mechanism of its action. Methods ·Overexpression of DUX cell lines in mESCs was achieved by using a lentiviral system. The proportion of 2-cell-like cells (2CLCs) before and after DUX overexpression was detected by flow cytometry, and the expression of 2-cell stage-specific genes, Dux, zinc finger and SCAN domain containing 4c (Zscan4c), zinc finger protein 352 (Zfp352) and murine endogenous retrovirus-L polymerase (MERVL-pol), were detected by real-time quantitative reverse transcription polymerase chain reaction (RT-qPCR). RT-qPCR assay was used to detect the expression of pluripotency factors, nanog homeobox (Nanog), kruppel-like transcription factor 4 (Klf4), sex determining region Y-box 2 (Sox2), and octamer-binding transcription factor 4 (Oct4), in pluripotent state, as well as the expression of signature genes for different germ layers in the differentiated state [endodermal: GATA binding protein 4 (Gata4), GATA binding protein 6 (Gata6), and sex determining region Y-box 17 (Sox17); ectodermal: Nestin and tubulin beta 3 class Ⅲ (Tubb3); mesodermal: heart and neural crest derivatives expressed 1 (Hand1), myogenic differentiation 1 (Myod1), and kinase insert domain protein receptor (Flk1)]. Public RNA sequencing (RNA-seq) data were mined to further clarify the effect of DUX on the differentiation of mESCs into extraembryonic endoderm. Functional and pathway enrichment analyses of differentially expressed genes were performed using Gene Ontology (GO), Kyoto Encyclopedia of Genes and Genomes (KEGG), and gene set enrichment analysis (GSEA) to identify the signaling pathways regulated by DUX. Additionally, an in-depth analysis of existing chromatin immunoprecipitation sequencing (ChIP-seq) data was conducted to explore the potential target genes of DUX. Results ·Molecular biology experiments showed that overexpression of DUX could effectively maintain the pluripotency of mESCs, which was consistent with the analysis of public RNA-seq data. Differential gene analysis revealed that endodermal genes were specifically upregulated. After differentiation assay of mESCs, RT-qPCR assay experiments showed that mRNA expression of the XEN marker genes (Gata4, Gata6, Sox17) was significantly upregulated (P<0.001). In contrast, there was no specific change in mesodermal and ectodermal genes. GSEA enrichment analysis indicated that DUX might activate the retinoid metabolism signaling pathway, and the analysis of the ChIP-seq data further revealed the presence of a large number of known retinoic acid receptor motif in DUX-bound peaks, which could activate downstream target genes related to the development of the XEN. Conclusion ·DUX has a strong correlation with the retinoic acid signaling pathway and it is predicted to activate the retinoic acid signaling pathway, which could promote the tendency of mESCs toward XEN differentiation.
HONG Lei, GUO Chuanliang, CAI Qin, LI Wanrui, ZENG Yitao, XUE Yan, ZENG Fanyi. Mechanism of DUX-induced differentiation of mESC into extraembryonic endoderm. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2024, 44(11): 1359-1369 doi:10.3969/j.issn.1674-8115.2024.11.003
MEM培养基、胎牛血清、β-巯基乙醇、胰蛋白酶(Gibco,美国),青-链霉素双抗、L-谷氨酰胺、非必需氨基酸(上海源培生物科技股份有限公司),LIF(Millipore,美国),PD0325901、CHIR99021(上海陶术生物科技有限公司),明胶(Sigma,德国),Lipofectamine 3000、Trizol(Invitrogen,美国),潮霉素、嘌呤霉素、阿霉素、氨苄霉素、酵母粉、胰蛋白胨(上海碧云天生物有限公司),聚合酶链反应(polymerase chain reaction,PCR)高保真酶、SYBR Green Fast qPCR Mix、cDNA反转录试剂盒(南京诺唯赞生物科技股份有限公司),感受态、质粒提取试剂盒(天根生化科技有限公司),焦碳酸二乙酯(diethyl pyrocarbonate,DEPC)、无水乙醇、异丙醇、三氯甲烷(上海大合化学品有限公司)。PCR仪(Roche,瑞士),实时荧光定量PCR仪、Nanodrop 2000、细胞培养箱(ThermoFisher,美国),显微镜(Olympus,日本)。
为了分析基因表达,本文使用trim-galore(v0.5.0)对原始数据进行过滤,得到高质量的clean data。利用hisat2(v2.1.0)软件将质量控制后的序列比对到参考基因组(mm10),并采用stringtie2(v2.1.7)软件分析对已知的基因和转录本进行表达定量。通过DESeq2分析差异表达基因,筛选标准为P<0.05且差异倍数(fold change,FC)对数的绝对值(|log2FC|)>1。使用R(v4.3.1)包clusterProfile(v4.8.2)对筛选出的差异基因进行基因本体论(Gene Ontology,GO)富集分析、京都基因和基因组数据库(Kyoto Encyclopedia of Genes and Genomes,KEGG)富集分析和基因集富集分析(gene set enrichment analysis,GSEA)。
使用bowtie2(v2.2.5)将ChIP-seq reads比对到参考基因组(mm10)上,丢弃线粒体DNA或未分配对应序列的读取。对于单一的比对,使用SAMtools (v1.9)过滤重复读取和低映射质量读取。使用MACS2(v2.1.2)调取ChIP-seq富集峰,用IDR (v2.0.3)合并共同峰,用于后续分析。使用R包ChIPseeker(v1.39.0)对富集的峰进行注释。使用MEME套件v5.0.5中的analysis of motif enrichment (AME)软件进行转录因子结合位点富集分析,从JASPAR下载用于扫描的结合基序矩阵。使用deeptools(v3.5.5)和RPKM(reads per kilo base per million mapped reads)归一化方法生成bigwig文件。在IGV(Integrative Genomics Viewer)中创建并展示图表。
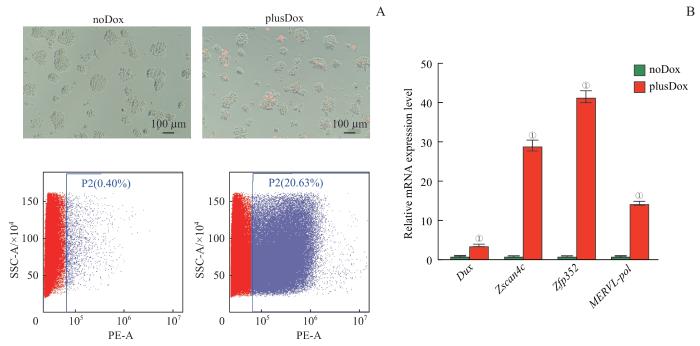
Note: A. Microscopy pictures and flow cytometry showing the percentage of 2CLCs. B. RT-qPCR validation of selected 2C marker genes and MERVL-polymerase gene (MERVL-pol) in noDox (control) and plusDox (DUX overexpression) ES cells. ①P<0.001, compared with the control group.
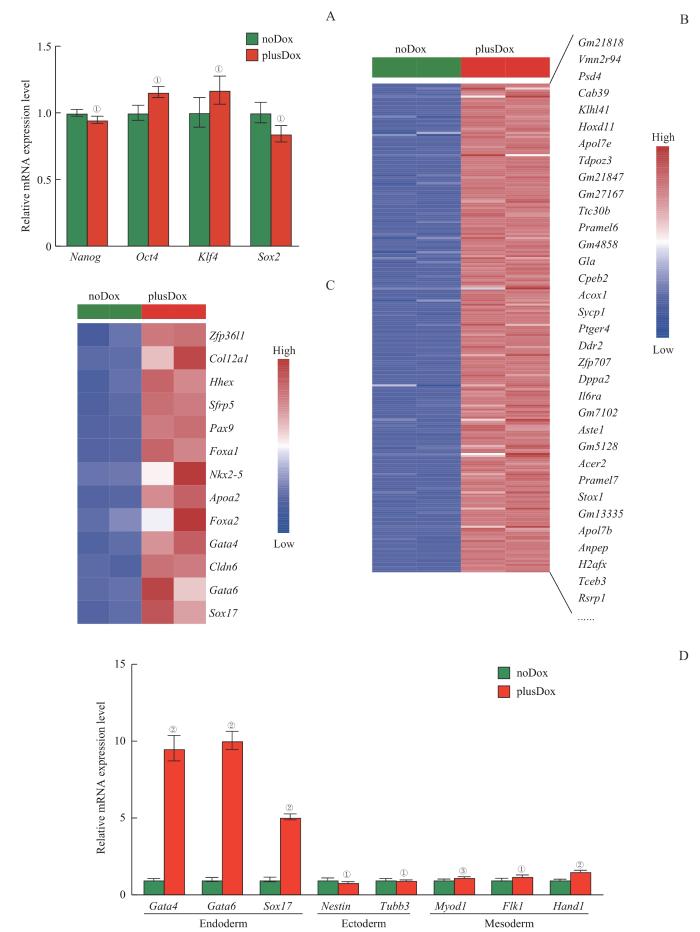
Note: A. RT-qPCR validation of selected pluripotent marker genes in noDox (control) and plusDox (DUX overexpression) ES cells. B/C. Heatmap representation of expression levels of genes involved in 2CLCs (B) or endoderm (C) specification in noDox (control) and plusDox (DUX overexpression) cells. D. RT-qPCR was employed to assess the RNA expression levels of marker factors for endoderm, mesoderm and ectoderm on the fourth day of embryoid body differentiation in noDox (control) and plusDox (DUX overexpression) cells. ①P>0.05, ②P<0.001, ③P=0.001 3, compared with the control group.
Fig 2
Detection of developmental potential of DUX-induced 2CLCs
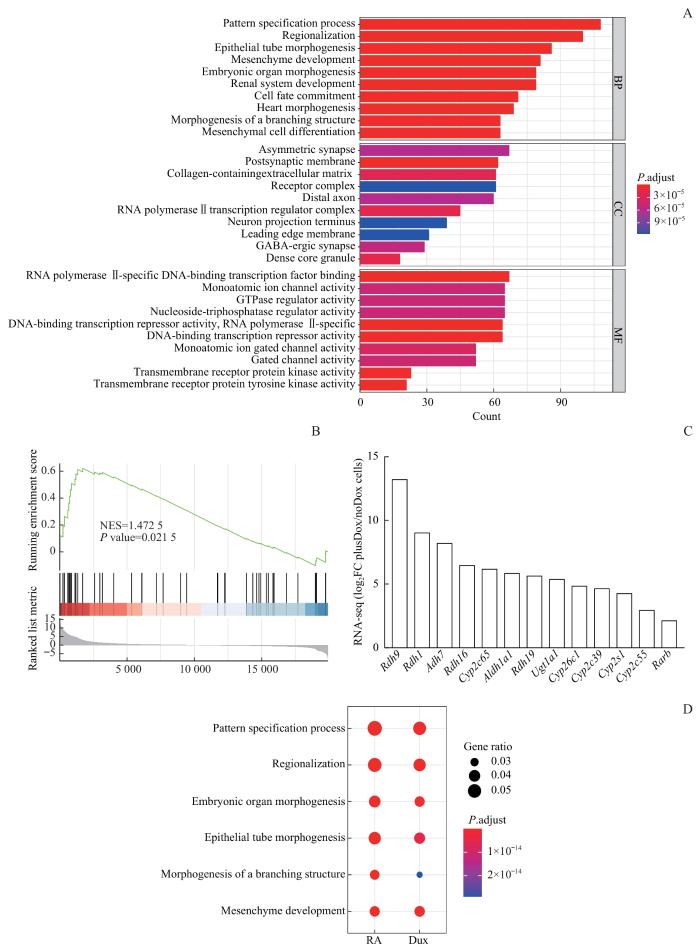
Note: A. GO analysis of upregulated genes in plusDox ES cells. B. GSEA analysis showing collective changes in the retinol metabolism signaling pathway gene set. C. Expression levels (log2FC) (FC, fold change) of selected RA-pathway-related genes in plusDox and noDox cells (ESCs) based on RNA-seq data. D. Comparison of enrichment results from multiple gene lists.
Fig 3
Functional enrichment analysis of DUX overexpressed ESCs
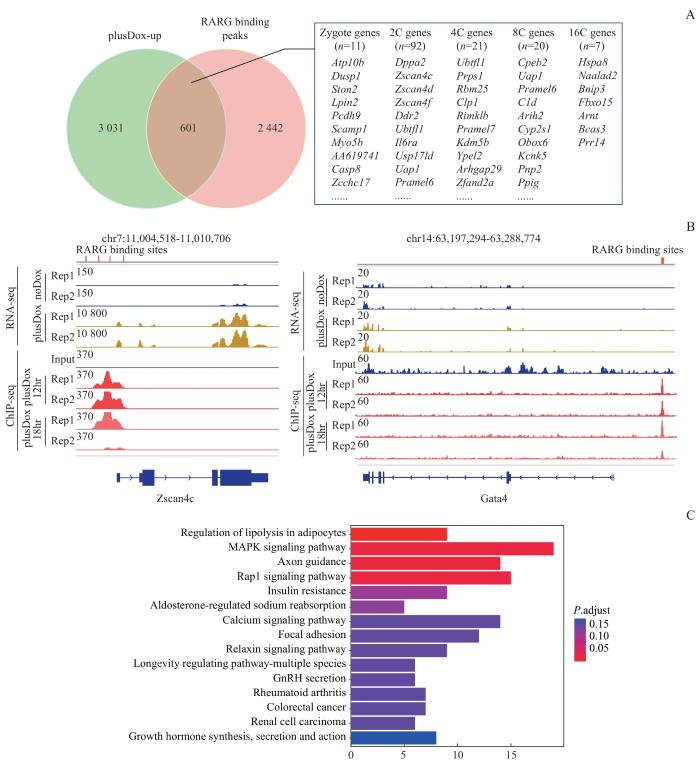
Note: A. Intersection of DUX-upregulated genes with RARG and DUX co-localized genes and possible mechanism of regulating mESCs. B. IGV view of RNA-seq and ChIP-seq results. C. KEGG analysis of the intersecting genes.
Fig 4
Potential mechanism of DUX regulation on ESCs differentiation
GSEA结果显示DUX激活了视黄醇代谢通路。视黄醇代谢产物视黄酸是核受体超家族的成员[25],控制着广泛的发育过程。最近有研究报道低剂量的视黄酸和视黄酸受体γ协同作用会促进mESC进入类2C状态[26]。本研究发现视黄酸途径促进的上调差异基因GO富集分析的功能区与DUX激活的高度相似,两者有相似的转录水平表达谱。这表明DUX可能通过视黄酸信号通路调节mESC状态。进一步对ChIP-seq数据进行生物信息学分析,证明已知的视黄酸受体motif存在于DUX富集的peaks中,两者能够结合在一些靶基因的启动子或增强子区域,调控这些基因的表达。有趣的是,RARG与DUX共同占据的基因,进一步与DUX激活的基因取交集,这些交集基因会富集在MAPK信号通路上,而持续激活的细胞外信号调节激酶1/2(extracellular regulated protein kinases 1/2,ERK1/2)磷酸化会导致mESC的分化[27]。此外,视黄酸信号通路也可直接激活MAPK/ERK通路[28],且视黄酸能够通过诱导GATA4等促分化因子的转录,促进内胚层来源的细胞分化[29]。本研究发现DUX与视黄酸有相似的结合基序,进而调控相同的靶基因,过表达DUX激活了视黄酸信号通路;如果抑制视黄酸受体,可能会抑制DUX与视黄酸下游相同的靶基因的转录,这些基因既有调节重编程相关的功能,也有促进分化的能力。
此外,本研究将为DUX异常激活导致的疾病治疗提供新思路。人类同源蛋白DUX4在成体肌肉细胞中的异常上调与面肩肱骨肌营养不良症(facioscapulohumeralmuscular dystrophy,FSHD)相关[30]。尽管DUX与DUX4的同源结构域和结合序列存在差异,但是它们都会激活2C基因的表达,其中可能存在的机制为两者通过结合并激活不同的内源性反转录转座子,导致临近基因的强烈表达[15]。在FSHD中DUX4的不当表达会引发细胞凋亡[31],而在DUX诱导的2CLC中也观察到严重的DNA损伤[32],这种细胞毒性可能源自2C时期特异性基因的异常激活,比如PRAME(preferentially expressed antigen in melanoma)家族基因[33]等。而在胚胎发育卵裂过程中,DUX的短暂激活并没有导致细胞死亡,这可能得益于ZSCAN4等相关因子的作用。已知ZSCAN4参与了ESC端粒和基因组稳定性的维持,以及保护2C胚胎免受DNA损伤[34-36]。因此,DUX增强了Zscan4的转录激活,会在一定程度上弥补其对细胞造成的伤害。在视黄酸处理的2CLC形成体系中,PRAME和ZSCAN4家族基因都会被激活[37-38]。鉴于DUX与视黄酸受体可能存在的交互影响,抑制视黄酸途径的某些关键分子可能在FSHD中抵消DUX4的作用,该领域的研究有望为此类疾病的靶向治疗奠定基础。
AUTHOR's CONTRIBUTIONS
The work of data analysis and the manuscript writing were completed by HONG Lei. The experimental operation was completed by HONG Lei, CAI Qin, and LI Wanrui. The manuscript was drafted and revised by GUO Chuanliang, XUE Yan, ZENG Yitao, and ZENG Fanyi. All authors have read the final manuscript and approved the submission.
MARTIN G R. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells[J]. Proc Natl Acad Sci U S A, 1981, 78(12): 7634-7638.
YANG Y, LIU B, XU J, et al. Derivation of pluripotent stem cells with in vivo embryonic and extraembryonic potency[J]. Cell, 2017, 169(2): 243-257.e25.
GAO X F, NOWAK-IMIALEK M, CHEN X, et al. Establishment of porcine and human expanded potential stem cells[J]. Nat Cell Biol, 2019, 21(6): 687-699.
LI R H, ZHONG C Q, YU Y, et al. Generation of blastocyst-like structures from mouse embryonic and adult cell cultures[J]. Cell, 2019, 179(3): 687-702.e18.
LIU K S, XU X C, BAI D D, et al. Bilineage embryo-like structure from EPS cells can produce live mice with tetraploid trophectoderm[J]. Protein Cell, 2023, 14(4): 262-278.
BOŠKOVIĆ A, EID A, PONTABRY J, et al. Higher chromatin mobility supports totipotency and precedes pluripotency in vivo[J]. Genes Dev, 2014, 28(10): 1042-1047.
SHEN H, YANG M, LI S Y, et al. Mouse totipotent stem cells captured and maintained through spliceosomal repression[J]. Cell, 2021, 184(11): 2843-2859.e20.
HENDRICKSON P G, DORÁIS J A, GROW E J, et al. Conserved roles of mouse DUX and human DUX4 in activating cleavage-stage genes and MERVL/HERVL retrotransposons[J]. Nat Genet, 2017, 49(6): 925-934.
DE IACO A, PLANET E, COLUCCIO A, et al. DUX-family transcription factors regulate zygotic genome activation in placental mammals[J]. Nat Genet, 2017, 49(6): 941-945.
FU X D, WU X J, DJEKIDEL M N, et al. Myc and Dnmt1 impede the pluripotent to totipotent state transition in embryonic stem cells[J]. Nat Cell Biol, 2019, 21(7): 835-844.
YANG F, HUANG X, ZANG R G, et al. DUX-miR-344-ZMYM2-mediated activation of MERVL LTRs induces a totipotent 2C-like state[J]. Cell Stem Cell, 2020, 26(2): 234-250.e7.
YANG G, ZHANG L F, LIU W Q, et al. Dux-mediated corrections of aberrant H3K9ac during 2-cell genome activation optimize efficiency of somatic cell nuclear transfer[J]. Cell Stem Cell, 2021, 28(1): 150-163.e5.
XU R M, LI S, WU Q, et al. Stage-specific H3K9me3 occupancy ensures retrotransposon silencing in human pre-implantation embryos[J]. Cell Stem Cell, 2022, 29(7): 1051-1066.e8.
ZUO F F, JIANG J Y, FU H P, et al. A TRIM66/DAX1/Dux axis suppresses the totipotent 2-cell-like state in murine embryonic stem cells[J]. Cell Stem Cell, 2022, 29(6): 948-961.e6.
WANG Y Q, NA Q, LI X H, et al. Retinoic acid induces NELFA-mediated 2C-like state of mouse embryonic stem cells associates with epigenetic modifications and metabolic processes in chemically defined media[J]. Cell Prolif, 2021, 54(6): e13049.
ITURBIDE A, RUIZ TEJADA SEGURA M L, NOLL C, et al. Retinoic acid signaling is critical during the totipotency window in early mammalian development[J]. Nat Struct Mol Biol, 2021, 28(6): 521-532.
YEN A, ROBERSON M S, VARVAYANIS S, et al. Retinoic acid induced mitogen-activated protein (MAP)/extracellular signal-regulated kinase (ERK) kinase-dependent MAP kinase activation needed to elicit HL-60 cell differentiation and growth arrest[J]. Cancer Res, 1998, 58(14): 3163-3172.
ARCECI R J, KING A A, SIMON M C, et al. Mouse GATA-4: a retinoic acid-inducible GATA-binding transcription factor expressed in endodermally derived tissues and heart[J]. Mol Cell Biol, 1993, 13(4): 2235-2246.
GENG L N, YAO Z Z, SNIDER L, et al. DUX4 activates germline genes, retroelements, and immune mediators: implications for facioscapulohumeral dystrophy[J]. Dev Cell, 2012, 22(1): 38-51.
MARKIEWICZ-POTOCZNY M, LOBANOVA A, LOEB A M, et al. TRF2-mediated telomere protection is dispensable in pluripotent stem cells[J]. Nature, 2021, 589(7840): 110-115.
SRINIVASAN R, NADY N, ARORA N, et al. Zscan4 binds nucleosomal microsatellite DNA and protects mouse two-cell embryos from DNA damage[J]. Sci Adv, 2020, 6(12): eaaz9115.
ZALZMAN M, FALCO G, SHAROVA L V, et al. Zscan4 regulates telomere elongation and genomic stability in ES cells[J]. Nature, 2010, 464(7290): 858-863.
TAGLIAFERRI D, MAZZONE P, NOVIELLO T M R, et al. Retinoic acid induces embryonic stem cells (ESCs) transition to 2 cell-like state through a coordinated expression of Dux and Duxbl1[J]. Front Cell Dev Biol, 2019, 7: 385.
NAPOLITANO G, TAGLIAFERRI D, FUSCO S, et al. A novel member of Prame family, Gm12794c, counteracts retinoic acid differentiation through the methyltransferase activity of PRC2[J]. Cell Death Differ, 2020, 27(1): 345-362.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}