Functional site analysis of mucin 1 in regulating the malignant characteristics of tumor cells
GAO Kexing,, LIAO Chunhua, LI Shengze, MA Shuangyu, HUANG Lei,
Department of Histoembryology, Genetics and Developmental Biology, Shanghai Jiao Tong University College of Basic Medical Sciences, Shanghai 200025, China
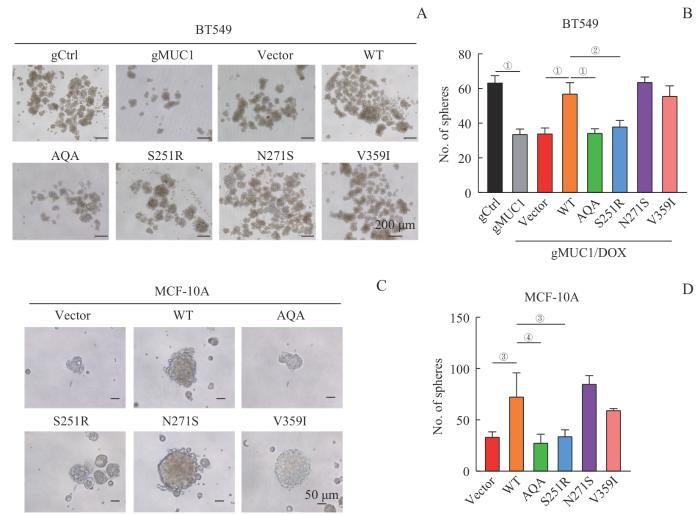
目的·探究黏蛋白1(mucin 1,MUC1)调控肿瘤细胞增殖、迁移和干性维持的功能位点。方法·通过癌症基因组图谱(The Cancer Genome Atlas,TCGA)数据库分析寻找MUC1基因在不同癌症中的突变特征,对不同MUC1突变位点进行分析及定位,并按突变出现频率排序;通过Western blotting筛选出突变频率较高且蛋白稳定表达的MUC1突变体,利用乳腺癌细胞株BT549敲除MUC1细胞系和乳腺非转化细胞株MCF-10A,应用慢病毒表达系统构建MUC1野生型(MUC1-WT)和突变体稳定表达的细胞系。采用免疫荧光法检测不同MUC1突变体的细胞定位。以MUC1-WT为阳性对照、MUC1-AQA功能丧失突变体为阴性对照,对不同突变细胞的肿瘤生物学功能进行分析:通过细胞计数试剂盒-8(cell counting kit-8,CCK-8)及克隆形成实验检测细胞增殖能力;通过划痕实验及Transwell实验检测细胞迁移能力;通过成球实验检测细胞干性。使用PyMOL软件分析MUC1突变体结构定位并通过蛋白质对接软件(ZDOCK Server)进行分子对接分析。结果·在TCGA数据库中得到102个位于MUC1编码区的突变,其中P418S、S251R、V359I、N271S、N465H 5个错义突变出现频率较高且位于非数目可变串联重复序列(non-variable number of tandem repeats,non-VNTR)区域。进一步检测发现MUC1-S251R、N271S、V359I突变体可稳定表达;细胞定位分析发现这3个突变体主要分布于细胞质,同时细胞核也有一定的分布,核质比与野生型未见明显差异。表达不同MUC1突变体细胞的肿瘤生物学功能分析发现:① MUC1-WT高表达显著增强BT549和MCF-10A细胞的增殖能力;与MUC1-WT细胞相比,MUC1-AQA、S251R、N271S突变体细胞增殖能力下降,但MUC1-V359I突变体细胞与MUC1-WT细胞具有相似的增殖能力。② MUC1-WT高表达细胞的迁移能力显著增强,而MUC1-AQA细胞迁移能力减弱。在BT549细胞中,MUC1-S251R与MUC1-V359I突变体细胞迁移能力与MUC1-WT细胞相似,但MUC1-N271S细胞的迁移能力较MUC1-WT细胞降低。在MCF-10A细胞中,MUC1-N271S与MUC1-V359I细胞的迁移能力接近MUC1-WT细胞;但MUC1-S251R细胞较MUC1-WT细胞迁移能力显著下降。③ MUC1-WT高表达显著增强2种细胞的干性,而MUC1-AQA细胞干性丧失;MUC1-N271S、V359I与MUC1-WT具有相似的维持细胞干性的能力,而MUC1-S251R使细胞干性减弱。PyMOL软件分析结果显示,MUC1-N271S及V359I位于海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)区域及附近,分别处于loop区及β-折叠处;分子对接结果显示,MUC1-WT及V359I与表皮生长因子受体(epidermal growth factor receptor,EGFR)胞外域形成复合物的稳定性强于MUC1-N271S和S251R,其稳定性排序为V359I>WT>N271S>S251R。结论·MUC1突变体对肿瘤细胞的生物学功能具有不同影响,其对增殖能力影响可能与EGFR信号通路相关。MUC1-V359I与MUC1-WT相似,并未影响MUC1对肿瘤细胞增殖、迁移及干性维持的作用;而MUC1-S251、N271位点可能参与细胞增殖和迁移的信号通路调控且MUC1-S251位点对维持细胞干性较为重要。
关键词:黏蛋白1
;
错义突变
;
肿瘤
;
细胞增殖
;
细胞迁移
;
细胞干性
Abstract
Objective ·To identify the functional motifs of mucin 1 (MUC1) involved in regulating tumor cell proliferation, migration, and stemness maintenance. Methods ·Mutational characteristics of the MUC1 gene across different cancers were identified from The Cancer Genome Atlas (TCGA) database. Various MUC1 mutation sites were analyzed and localized, followed by ranking based on mutation frequency. Western blotting was used to screen high-frequency MUC1 mutants with stable protein expression. BT549 cell line with MUC1 knocked out and MCF-10A cell line were used to stably overexpress MUC1 wild-type (MUC1-WT) and mutants by using lentiviral technology. Immunofluorescence was used to detect the cellular localization of MUC1 mutants. Using MUC1-WT as a positive control and MUC1-AQA, a loss-of-function mutant, as a negative control, the biological functions of different MUC1 mutant cells were analyzed: cell proliferation ability was assessed by cell counting kit-8 (CCK-8) assay and colony formation assay; cell migration ability was evaluated by wound-healing and Transwell assays; cell stemness was examined by sphere formation assay. Structural localization of MUC1 mutants was analyzed by using PyMOL software, and molecular docking analysis was performed by using a protein docking software (ZDOCK Server). ·Results A total of 102 mutations located in the MUC1 coding region were identified in the TCGA database, among which five missense mutations (P418S, S251R, V359I, N271S, and N465H) exhibited higher frequencies and were located in the non-variable number of tandem repeats (non-VNTR) region. Further examination revealed that the MUC1-S251R, N271S, and V359I mutants could be stably expressed. The cellular localization assay indicated that these three mutants predominantly localized in the cytoplasm, but were also presented in the nucleus. The nuclear-to-cytoplasmic ratio showed minimal differences between MUC1-WT and the mutants. Analysis of the tumorigenic biological functions of the cells expressing different MUC1 mutants revealed that: ① High expression of MUC1-WT significantly enhanced the proliferation ability of both BT549 and MCF-10A cells; the proliferation of MUC1-AQA, S251R, and N271S mutant cells was decreased compared to MUC1-WT cells, while MUC1-V359I mutant cells exhibited a similar proliferative profile to MUC1-WT cells. ② The migration ability of MUC1-WT high-expressing cells was significantly enhanced, whereas MUC1-AQA cells demonstrated attenuated migration. In the BT549 cells, the migration ability of MUC1-S251R and V359I cells was similar to that of MUC1-WT cells, whereas MUC1-N271S cells showed reduced migration. In the MCF-10A cells, the migration ability of MUC1-N271S and MUC1-V359I cells was similar to that of MUC1-WT cells, whereas MUC1-S251R cells exhibited significantly decreased migration. ③ Stemness was enhanced in both cell types with high MUC1-WT expression, while MUC1-AQA cells lost stemness; the cells with MUC1-N271S, V359I and MUC1-WT showed comparable maintenance of stemness, whereas MUC1-S251R cells demonstrated compromised stemness. PyMOL software analysis unveiled that MUC1-N271S and V359I were located in or around the sperm protein-enterokinase-agarin (SEA) region, specifically in the loop region and the β-sheet, respectively. The molecular docking analysis revealed that the stability of the complex formed by MUC1-WT or V359I with the extracellular domain of epidermal growth factor receptor (EGFR) surpassed that of MUC1-N271S or S251R, indicating a stability hierarchy of V359I>WT>N271S>S251R. ·Conclusion MUC1 mutants exhibit diverse impacts on the biological functions of tumor cells, with their effects on proliferation correlating with the EGFR signaling pathway. MUC1-V359I is similar to MUC1-WT, indicating a negligible effect on tumor cell proliferation, migration, and stemness maintenance. Conversely, MUC1-S251 and N271 sites may be involved in the regulation of signaling pathways governing cell proliferation and migration and the MUC1-S251 site plays a critical role in maintaining cell stemness.
GAO Kexing, LIAO Chunhua, LI Shengze, MA Shuangyu, HUANG Lei. Functional site analysis of mucin 1 in regulating the malignant characteristics of tumor cells. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2024, 44(11): 1370-1382 doi:10.3969/j.issn.1674-8115.2024.11.004
黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2]。MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6]。MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8]。MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10]。MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11]。MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14]。MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面。正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17]。此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18]。因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物。
MUC1,特别是MUC1-C,在肿瘤中参与多种信号通路的异常转导,调控肿瘤细胞增殖、转移、凋亡、干性维持及化学治疗耐药等恶性特征[21],但是MUC1参与肿瘤各种恶性特征调控的基序并不完全清楚。过往研究主要聚焦于MUC1过表达与肿瘤的关系,关于MUC1的基因突变研究仅限于由VNTR区AG缺失导致的移码突变,进而引发的常染色体显性遗传肾小管间质性肾病(autosomal dominant tubulointerstitial kidney disease,ADTKD)[22-23]。目前尚无研究对肿瘤中MUC1的错义突变及功能进行分析。为了研究MUC1蛋白参与肿瘤恶性特征调控的功能基序,我们通过癌症基因组图谱(The Cancer Genome Atlas,TCGA)数据库分析,寻找MUC1在不同癌症中的突变位点;并选择突变频率较高且稳定表达的突变体,研究它们对肿瘤细胞增殖、迁移、干性维持的影响,以探究MUC1不同位点的功能。
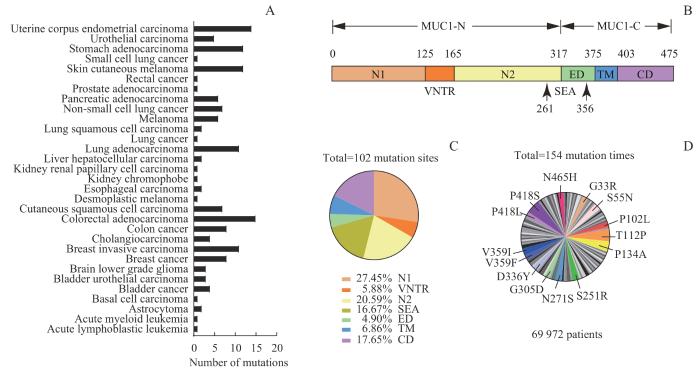
Note: A. Proportion of MUC1 mutations in various tumor types. B. MUC1 protein consisting of N1, VNTR, N2, SEA, ED, TM, and CD domains. C. Location of 102 mutation sites in MUC1 protein. D. The mutation sites with a frequency of 1.95% or more in tumor samples.
Fig 1
Analysis of sources, positions, and frequencies of MUC1 mutation sites in the TCGA database
Tab 2
表2
表214个 MUC1 高频突变体的信息
Tab 2 Information of 14 MUC1 mutants with high frequency
Note: A/C/E. Western blotting was used to detect MUC1 protein expression levels in HEK293T cells (A), BT549 cells (C) and MCF-10A cells (E). B/D/F. Statistical analysis of MUC1 protein levels in HEK293T cells (B), BT549 cells (D) and MCF-10A cells (F). ①P<0.001, ②P=0.020, ③P=0.002.
Fig 2
Protein expression levels of MUC1-WT and mutants in the cells
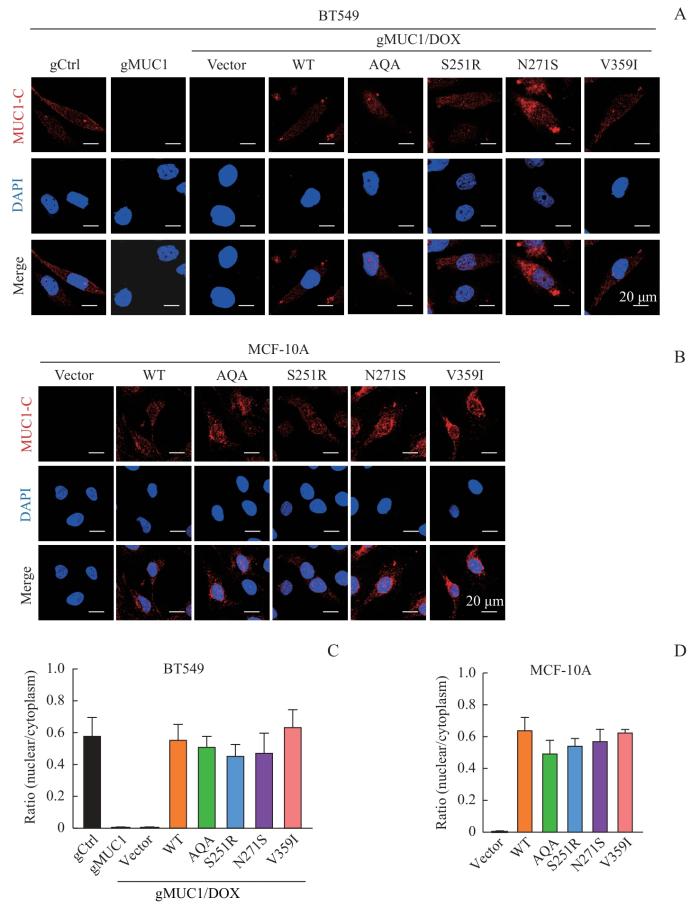
Note: A/B. Detection of cellular localization of MUC1 mutants by immunofluorescence in BT549 cells (A) and MCF-10A cells (B)(×600). C/D. Statistical analysis of MUC1 nuclear/cytoplasm ratio in BT549 cells (C) and MCF-10A cells (D).
Fig 3
Cellular localization of MUC1-WT and mutants
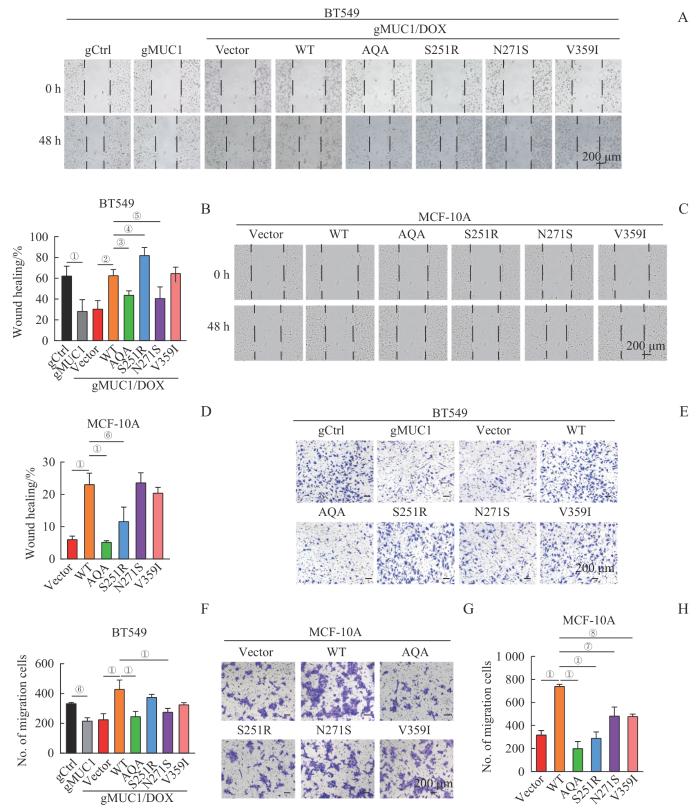
Note: A/B. Wound healing assay of BT549 cells expressing MUC1-WT or mutants. Representative images of cells at 0 h and 48 h (A, ×200) and corresponding statistical analysis (B). C/D. Wound healing assay of MCF-10A cells expressing MUC1-WT or mutants. Representative images of cells at 0 h and 48 h (C, ×200) and corresponding statistical analysis (D). E/F. Transwell migration analysis of BT549 cells expressing MUC1-WT or mutants. Representative crystal violet-stained images (E, ×100) and corresponding statistical analysis (F). G/H. Transwell migration analysis of MCF-10A cells expressing MUC1-WT or mutants. Representative crystal violet-stained images (G, ×100) and corresponding statistical analysis (H). ①P<0.001, ②P=0.004, ③P=0.014, ④P=0.030, ⑤P=0.025, ⑥P=0.003, ⑦P=0.050, ⑧P=0.042.
Fig 5
Effect of MUC1-WT and mutants on cell migration
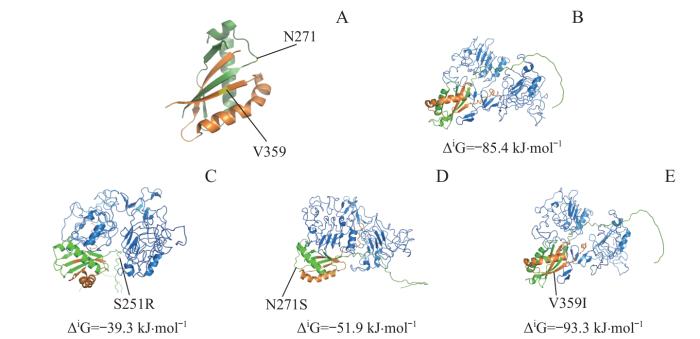
Note: A. PyMoL software was used to localize MUC1 mutants in or around the SEA domain. B‒E. ZDOCK Server was used to analyze the molecular docking of MUC1-WT (B) and mutants (C, S251R; D, N271S; E, V359I) with EGFR-ECD. Green represents the MUC1-N region (before the GSVVV sequence), orange represents the MUC1-C region (after the GSVVV sequence), blue represents EGFR-ECD region, and yellow represents the localization of the mutation sites.
Fig 7
Analysis of localization of MUC1 mutation sites in or around the SEA domain and molecular docking of the mutants with EGFR-ECD
The experimental operation and data analysis were completed by GAO Kexing, LIAO Chunhua and LI Shengze. GAO Kexing, MA Shuangyu and HUANG Lei were responsible for manuscript draft and revision. HUANG Lei was responsible for project design. All authors have read the final manuscript and agreed to the submission.
利益冲突声明
所有作者声明不存在利益冲突
COMPETING INTERESTS
All authors disclose no relevant conflict of interests
ANDRIANIFAHANANA M, MONIAUX N, BATRA S K. Regulation of mucin expression: mechanistic aspects and implications for cancer and inflammatory diseases[J]. Biochim Biophys Acta, 2006, 1765(2): 189-222.
MONIAUX N, ANDRIANIFAHANANA M, BRAND R E, et al. Multiple roles of mucins in pancreatic cancer, a lethal and challenging malignancy[J]. Br J Cancer, 2004, 91(9): 1633-1638.
GAEMERS I C, VOS H L, VOLDERS H H, et al. A STAT-responsive element in the promoter of the episialin/MUC1 gene is involved in its overexpression in carcinoma cells[J]. J Biol Chem, 2001, 276(9): 6191-6199.
SHENG Y H, LOURIE R, LINDÉN S K, et al. The MUC13 cell-surface mucin protects against intestinal inflammation by inhibiting epithelial cell apoptosis[J]. Gut, 2011, 60(12): 1661-1670.
SINGH A P, MONIAUX N, CHAUHAN S C, et al. Inhibition of MUC4 expression suppresses pancreatic tumor cell growth and metastasis[J]. Cancer Res, 2004, 64(2): 622-630.
RAINA D, AHMAD R, RAJABI H, et al. Targeting cysteine-mediated dimerization of the MUC1-C oncoprotein in human cancer cells[J]. Int J Oncol, 2012, 40(5): 1643-1649.
VAN PUTTEN J P M, STRIJBIS K. Transmembrane mucins: signaling receptors at the intersection of inflammation and cancer[J]. J Innate Immun, 2017, 9(3): 281-299.
YOLKEN R H, PETERSON J A, VONDERFECHT S L, et al. Human milk mucin inhibits rotavirus replication and prevents experimental gastroenteritis[J]. J Clin Invest, 1992, 90(5): 1984-1991.
LAU S K, WEISS L M, CHU P G. Differential expression of MUC1, MUC2, and MUC5AC in carcinomas of various sites: an immunohistochemical study[J]. Am J Clin Pathol, 2004, 122(1): 61-69.
SAFI F, KOHLER I, RÖTTINGER E, et al. The value of the tumor marker CA 15-3 in diagnosing and monitoring breast cancer[J]. Cancer, 1991, 68(3): 574-582.
YAMAMOTO S, KAIMORI J Y, YOSHIMURA T, et al. Analysis of an ADTKD family with a novel frameshift mutation in MUC1 reveals characteristic features of mutant MUC1 protein[J]. Nephrol Dial Transplant, 2017, 32(12): 2010-2017.
WENZEL A, ALTMUELLER J, EKICI A B, et al. Single molecule real time sequencing in ADTKD-MUC1 allows complete assembly of the VNTR and exact positioning of causative mutations[J]. Sci Rep, 2018, 8(1): 4170.
LI Q F, CHU Y K, LI S Z, et al. The oncoprotein MUC1 facilitates breast cancer progression by promoting Pink1-dependent mitophagy via ATAD3A destabilization[J]. Cell Death Dis, 2022, 13(10): 899.
JIN W, LIAO X D, LV Y P, et al. MUC1 induces acquired chemoresistance by upregulating ABCB1 in EGFR-dependent manner[J]. Cell Death Dis, 2017, 8(8): e2980.
RAZAWI H, KINLOUGH C L, STAUBACH S, et al. Evidence for core 2 to core 1 O-glycan remodeling during the recycling of MUC1[J]. Glycobiology, 2013, 23(8): 935-945.
PASTRELLO C, SANTAROSA M, FORNASARIG M, et al. MUC gene abnormalities in sporadic and hereditary mucinous colon cancers with microsatellite instability[J]. Dis Markers, 2005, 21(3): 121-126.
ZHANG L X, VLAD A, MILCAREK C, et al. Human mucin MUC1 RNA undergoes different types of alternative splicing resulting in multiple isoforms[J]. Cancer Immunol Immunother, 2013, 62(3): 423-435.
LIAO C H, YU L P, PANG Z, et al. WWP1 targeting MUC1 for ubiquitin-mediated lysosomal degradation to suppress carcinogenesis[J]. Signal Transduct Target Ther, 2021, 6(1): 297.
ZHANG W B, LIU M W, YU L L, et al. Perturbation effect of single polar group substitution on the self-association of amphiphilic peptide helices[J]. J Colloid Interface Sci, 2022, 610: 1005-1014.
ZHANG W B, LIU M W, WANG Y, et al. β-sheet assembly translates conservative single-site mutation into a perturbation in macroscopic structure[J]. Nano Lett, 2023, 23(6): 2370-2378.
MACAO B, JOHANSSON D G, HANSSON G C, et al. Autoproteolysis coupled to protein folding in the SEA domain of the membrane-bound MUC1 mucin[J]. Nat Struct Mol Biol, 2006, 13(1): 71-76.
MERLIN J, STECHLY L, DE BEAUCÉ S, et al. Galectin-3 regulates MUC1 and EGFR cellular distribution and EGFR downstream pathways in pancreatic cancer cells[J]. Oncogene, 2011, 30(22): 2514-2525.
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
1
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
1
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
1
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
1
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
1
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
2
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
1
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
1
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
1
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
1
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
1
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
1
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
1
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
1
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
1
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
1
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
1
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
1
... 黏蛋白(mucin,MUC)家族是一类大分子糖蛋白,有21个家族成员,可分为跨膜型和分泌型2种类型[1-2].MUC家族多个成员的异常表达与癌症和炎症性疾病相关,例如MUC13、MUC17的表达上调与胃癌和胰腺癌的发生有关[3-4],MUC13基因敲除可以使肠上皮细胞凋亡增加从而促进肠道炎症的发生[5-6].MUC1基因位于1号染色体q22区域,转录为单一的mRNA,翻译产生的肽链在内质网中被水解为N末端亚基(MUC1-N)和C末端亚基(MUC1-C),两者通过非共价键连接形成MUC1蛋白[7-8].MUC1-N包含数目可变串联重复序列(variable number of tandem repeats,VNTR)区和部分海胆精子蛋白-肠激酶-聚集蛋白(sperm protein-enterokinase-agarin,SEA)结构域;其中,VNTR区域富含丝氨酸(serine,Ser)、苏氨酸(threonine,Thr)和脯氨酸(proline,Pro),是发生O-连接N-乙酰半乳糖胺(GalNAc)修饰的位点;SEA结构域中的GSVVV基序是MUC1被水解为2个亚基的关键位点[9-10].MUC1-C由胞外段(extracellular domain,ED)、跨膜段(transmembrane domain,TM)及胞内段(cytoplasmic domain,CD)组成[11].MUC1-CD含有7个酪氨酸(tyrosine,Tyr),可以参与多种激酶信号通路[12];MUC1-CD头部含CQC(半胱氨酸-谷氨酰胺-半胱氨酸)基序,是MUC1形成同源二聚体并行使生物学功能所必需[13-14].MUC1在组织器官中广泛分布,主要包括消化道、生殖管道、呼吸系统,以及各种腺体的管腔上皮细胞表面.正常情况下,MUC1与MUC家族其他成员一起形成黏液层,润滑并保护上皮细胞免受环境因素的刺激[15-16];在肿瘤组织中,MUC1丧失极性分布并过度表达,其蛋白水平可达到正常水平的50~100倍[17].此外,MUC1侧链被高度唾液酸化,阻止糖链的进一步延伸,从而暴露配体结合域并增强致癌信号[18].因此,MUC1的胞外抗原CA15-3[19]和CA19-9[20]已作为乳腺癌和胰腺癌诊断的临床标志物. ...
1
... MUC1,特别是MUC1-C,在肿瘤中参与多种信号通路的异常转导,调控肿瘤细胞增殖、转移、凋亡、干性维持及化学治疗耐药等恶性特征[21],但是MUC1参与肿瘤各种恶性特征调控的基序并不完全清楚.过往研究主要聚焦于MUC1过表达与肿瘤的关系,关于MUC1的基因突变研究仅限于由VNTR区AG缺失导致的移码突变,进而引发的常染色体显性遗传肾小管间质性肾病(autosomal dominant tubulointerstitial kidney disease,ADTKD)[22-23].目前尚无研究对肿瘤中MUC1的错义突变及功能进行分析.为了研究MUC1蛋白参与肿瘤恶性特征调控的功能基序,我们通过癌症基因组图谱(The Cancer Genome Atlas,TCGA)数据库分析,寻找MUC1在不同癌症中的突变位点;并选择突变频率较高且稳定表达的突变体,研究它们对肿瘤细胞增殖、迁移、干性维持的影响,以探究MUC1不同位点的功能. ...
1
... MUC1,特别是MUC1-C,在肿瘤中参与多种信号通路的异常转导,调控肿瘤细胞增殖、转移、凋亡、干性维持及化学治疗耐药等恶性特征[21],但是MUC1参与肿瘤各种恶性特征调控的基序并不完全清楚.过往研究主要聚焦于MUC1过表达与肿瘤的关系,关于MUC1的基因突变研究仅限于由VNTR区AG缺失导致的移码突变,进而引发的常染色体显性遗传肾小管间质性肾病(autosomal dominant tubulointerstitial kidney disease,ADTKD)[22-23].目前尚无研究对肿瘤中MUC1的错义突变及功能进行分析.为了研究MUC1蛋白参与肿瘤恶性特征调控的功能基序,我们通过癌症基因组图谱(The Cancer Genome Atlas,TCGA)数据库分析,寻找MUC1在不同癌症中的突变位点;并选择突变频率较高且稳定表达的突变体,研究它们对肿瘤细胞增殖、迁移、干性维持的影响,以探究MUC1不同位点的功能. ...
1
... MUC1,特别是MUC1-C,在肿瘤中参与多种信号通路的异常转导,调控肿瘤细胞增殖、转移、凋亡、干性维持及化学治疗耐药等恶性特征[21],但是MUC1参与肿瘤各种恶性特征调控的基序并不完全清楚.过往研究主要聚焦于MUC1过表达与肿瘤的关系,关于MUC1的基因突变研究仅限于由VNTR区AG缺失导致的移码突变,进而引发的常染色体显性遗传肾小管间质性肾病(autosomal dominant tubulointerstitial kidney disease,ADTKD)[22-23].目前尚无研究对肿瘤中MUC1的错义突变及功能进行分析.为了研究MUC1蛋白参与肿瘤恶性特征调控的功能基序,我们通过癌症基因组图谱(The Cancer Genome Atlas,TCGA)数据库分析,寻找MUC1在不同癌症中的突变位点;并选择突变频率较高且稳定表达的突变体,研究它们对肿瘤细胞增殖、迁移、干性维持的影响,以探究MUC1不同位点的功能. ...





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}