目的·利用转录组以及脂质组分析技术研究瞬时受体电位香草素1型(transient receptor potential vanilloid type 1,TRPV1)通道的激活对高脂饮食诱导的小胶质细胞代谢的调控作用。方法·以8周龄C57BL/6J小鼠(WT)和Trpv1-/-(KO)小鼠为实验动物,高脂饲料(high-fat diet,HFD)分别喂养3d、7d、8周诱导造模(WT和KO组,n=3;WT-HFD和KO-HFD组,n=4)。通过免疫荧光试验测量WT-HFD和KO-HFD组小鼠大脑中TRPV1通道的表达以及细胞定位。通过RNA测序和液相色谱-质谱法确定WT-HFD和KO-HFD组小鼠的大脑表型。结果·与WT组小鼠相比,WT-HFD组小鼠体内小胶质细胞Trpv1 mRNA的表达水平显著增加。与WT-HFD组小鼠相比,KO-HFD组小鼠的脑脂质代谢、线粒体功能、葡萄糖转移以及糖酵解相关基因的表达水平下调。脂质组分析显示,虽然KO-HFD组小鼠的脑组织中脂质积累,但是Trpv1基因敲除减弱了HFD诱导的小胶质细胞活化,此外,TRPV1激动剂辣椒素在体外减弱棕榈酸诱导的线粒体膜电位去极化。结论·TRPV1通过线粒体驱动的燃料可用性机制调节小胶质细胞的脂质和葡萄糖代谢。
关键词:瞬时受体电位香草素1型
;
小胶质细胞
;
代谢
;
脂质
;
线粒体
Abstract
Objective ·Transcriptomic and lipidomic analysis techniques were used to investigate the role of transient receptor potential vanilloid type 1 (TRPV1) channel activation in the regulation of high-fat diet-induced microglial metabolism. Methods ·Eight-week-old C57BL/6J mice (WT) and Trpv1-/- (KO) mice were used as experimental animals, and fed high-fat diet (HFD) for 3 days, 7 days, and 8 weeks to induce modelling (WT and KO groups, n = 3; WT-HFD and KO-HFD groups, n = 4). TRPV1 channel expression and cellular localisation were measured by immunofluorescence in the brains of mice in the WT-HFD and KO-HFD group. RNA sequencing and liquid chromatography-mass spectrometry were performed to determine the brain phenotype of mice in the WT-HFD and KO-HFD groups. Results ·The expression level of Trpv1 mRNA in microglia was significantly increased in mice in the WT-HFD group compared to mice in the WT group. The expression levels of genes related to brain lipid metabolism, mitochondrial function, glucose transfer, and glycolysis were down-regulated in the KO-HFD group of mice compared with the WT-HFD group of mice. Lipidomic analysis showed that although lipids accumulated in the brain tissue of mice in the KO-HFD group, Trpv1 knockdown attenuated HFD-induced microglia activation, and in addition the TRPV1 agonist capsaicin attenuated palmitate-induced depolarisation of mitochondrial membrane potential in vitro. Conclusion ·Together, these findings suggest that TRPV1 regulates lipid and glucose metabolism in microglia via fuel availability driven by a mitochondrial mechanism.
SHA Xudong, WANG Chenfei, LU Jia, YU Zhihua. Regulation of high-fat diet-induced microglial metabolism by transient receptor potential vanilloid type 1. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2023, 43(12): 1493-1506 doi:10.3969/j.issn.1674-8115.2023.12.004
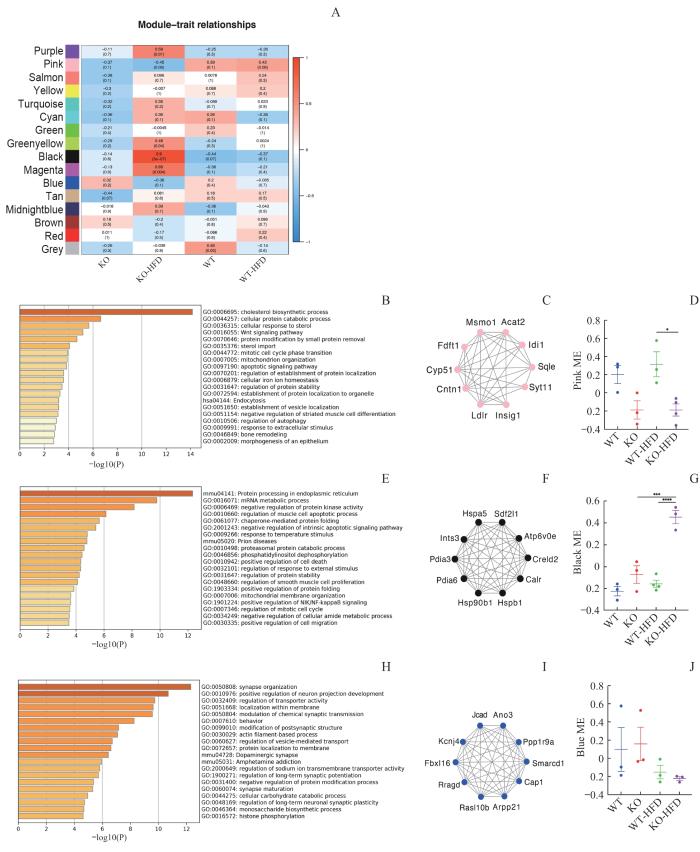
Note: A. Modules of WT, KO, WT-HFD, and KO-HFD mice (n=3 mice in WT and KO group, n=4 mice in WT-HFD and KO-HFD group). B/E/H. Top 20 pathways of GO and KEGG enrichment analysis of the pink, black and blue module. C/F/I. Network plot of the top 10 genes in pink, black and blue module. D/G/J. Trajectory of the module eigengenes in pink, black and blue module. One-way ANOVA was applied. *P < 0.05, ***P < 0.001, ****P = 0.000.
Fig 1
WGCNA in WT and genetic Trpv1 deletion mice before and after high fat feeding
Note: A. The composition of lipidome profiling of brain cells. B—C. Lipidome profiling of WT-HFD compared to WT and KO-HFD compared to KO mice brain. D. Distribution of PC, PE, and PG chain lengths of WT and KO mice induced by HFD treating. E—G. Distribution of PC, PE, and PG species of WT and KO mice induced by HFD treating. Data present the x±s (WT, WT-HFD, KO, n =3; KO-HFD, n =4). Statistical test: two-sided Student's t-test, two-way ANOVA followed by the Dunnett's post hoc test. *P < 0.05, **P < 0.01, ***P < 0.001, **** P = 0.000.
Fig 2
HFD induced liposome switch of brain cells with Trpv1 deficiency
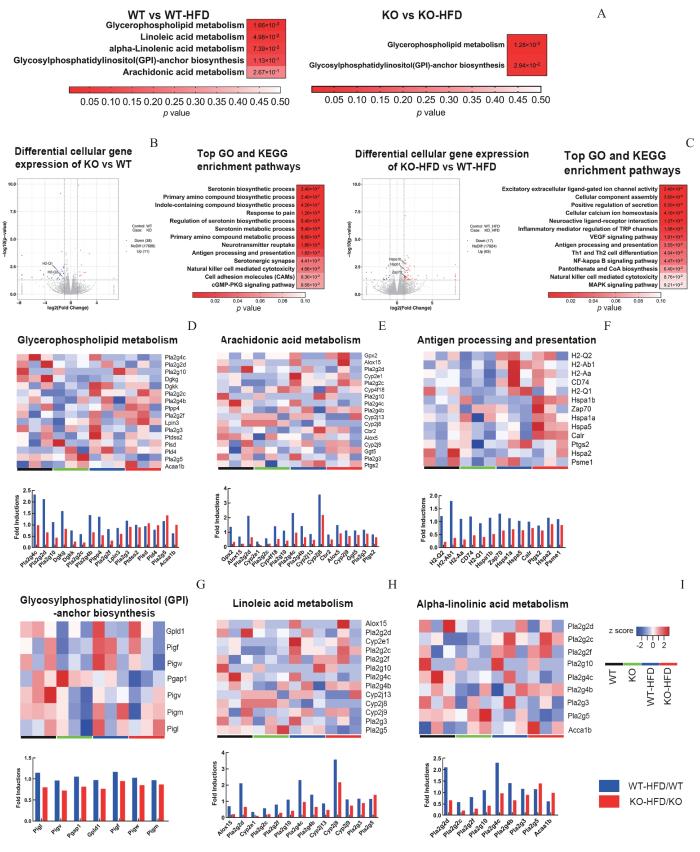
Note: A. Top lipidome enriched KEGG pathways of WT-HFD compared to WT mice and KO-HFD compared to KO mice. B/C. Gene expression changes, top GO and KEGG pathways enriched pathways of KO compared to WT mice and KO-HFD compared to WT-HFD mice.D—I. Heat maps of KO compared to WT mice and KO-HFD compared to WT-HFD mice.
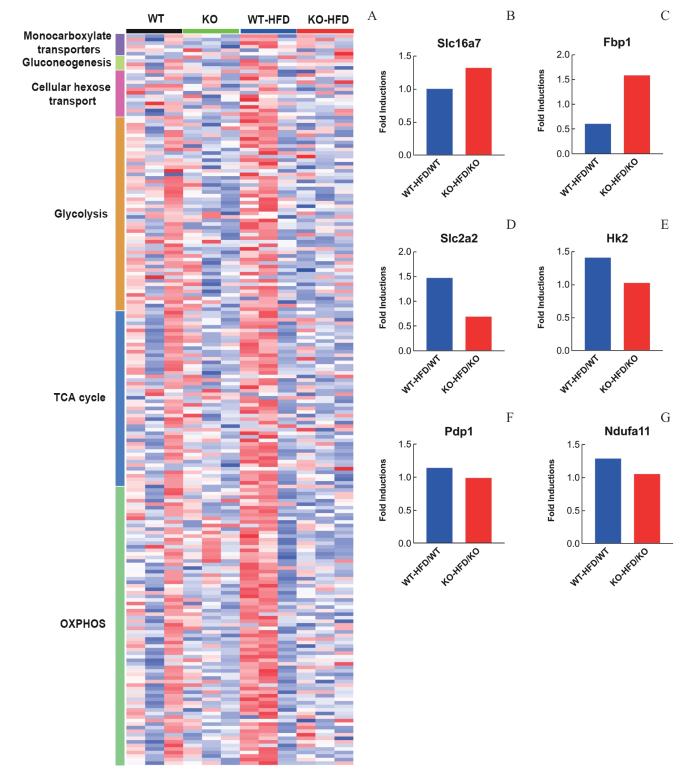
Note: A. The expressions of genes involved in monocarboxylate transporters, gluconeogenesis, glucose transporter, lactate shuttle, glycolysis, TCA cycle, and oxidative phosphorylation of WT and KO mice treated on SCD or 7 d HFD (red, increase; blue, decrease). B—G. The bar graphs show the fold induction of those genes within the class in WT-HFD versus WT mice (blue), and KO-HFD versus KO mice (red). All replicates within a class were averaged to obtain fold induction. Color key on the bottom of the figure indicated the group of samples.
Fig 4
Genetic Trpv1 deletion reduced HFD-induced multiple effects on metabolic pathways of the brain
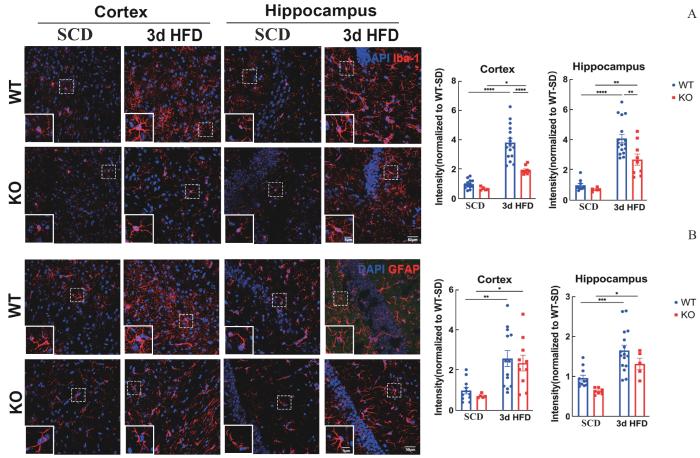
Note: A/B. Iba-1+ active microglia and GFAP+ reactive astrocytes of WT or TRPV1 KO mice fed on 3-day HFD. Data represent x±s. ⁎P < 0.05, **P < 0.01, ***P < 0.001, ****P = 0.000. Scale bar: 50 μm.
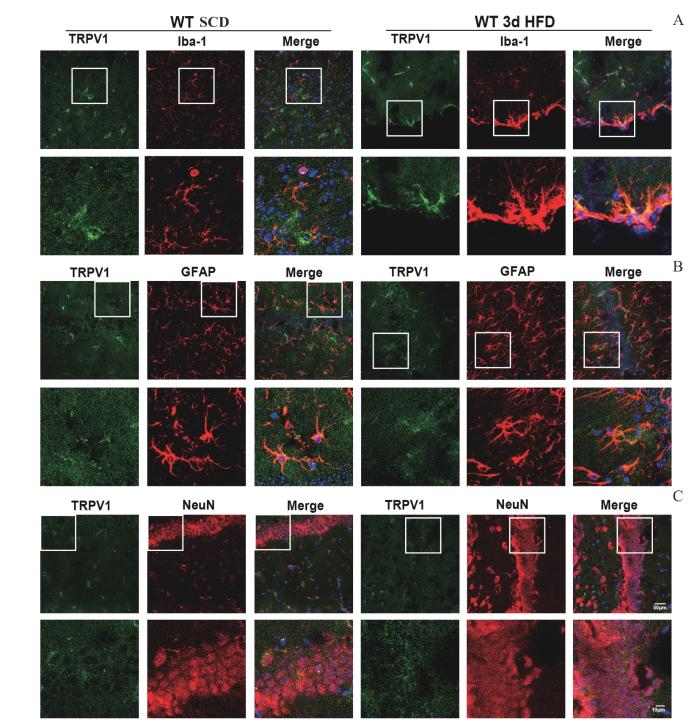
Note: A—C. Co-staining of TRPV1 and Iba1, TRPV1 and GFAP, TRPV1 and NeuN in SCD and 3-day HFD mice. Nuclei were stained in blue with DAPI. Scale bar: 50 μm; Scale bar: 10 μm.
Fig 6
Up-regulation of TRPV1 in active microglia and reactive astrocytes of HFD mice brains
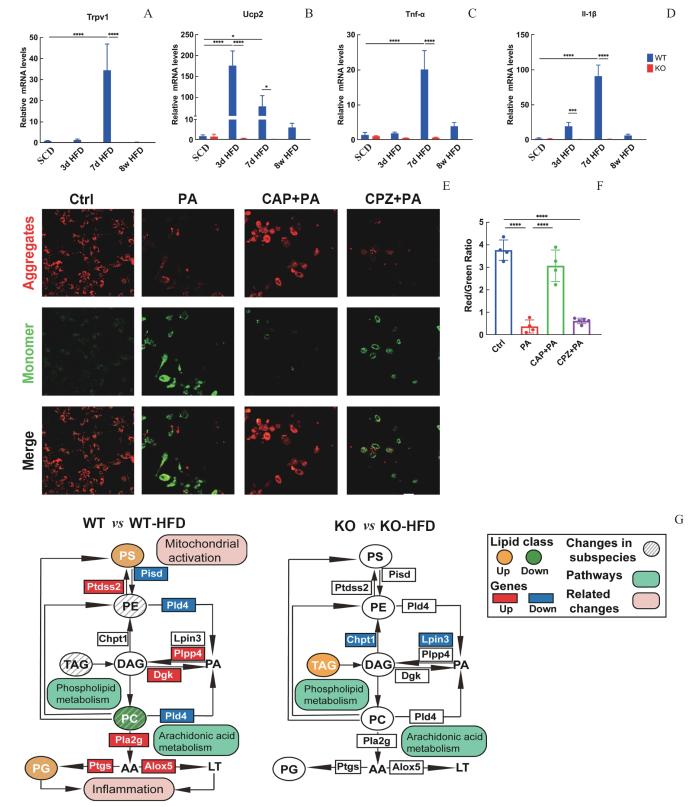
Note: A—D. The mRNA levels of Trpv1, Ucp2, Tnf-α, and Il-1β in isolated cortical microglia (CD11b+ cells) from WT and KO mice mixed gender fed on SCD, 3 d HFD, 7 d HFD, or 8-week HFD (n=3). E/F. Mitochondrial membrane potential was detected by fluorescence microscopy (n=3). G. Indications of quantified lipid classes and acyl chains (circles) and genes (rectangles) of WT and KO mice by HFD. Data represent x±s. ⁎P < 0.05, ****P = 0.000. Scale bar: 50 μm.
YU Zhihua was responsible for the experimental design; SHA Xudong participated in the experimental design and was responsible for data analysis and writing the paper. WANG Chenfei and LU Jia participated in the experimental design and were responsible for data analysis. All authors have read the final manuscript and agreed to the submission.
利益冲突声明
所有作者声明没有竞争性利益。
COMPETING INTERESTS
All authors declare that they have no competing interests.
HORVATH T L, SARMAN B, GARCÍA-CÁCERES C, et al. Synaptic input organization of the melanocortin system predicts diet-induced hypothalamic reactive gliosis and obesity[J]. Proc Natl Acad Sci USA, 2010, 107(33): 14875-14880.
VALDEARCOS M, DOUGLASS J D, ROBBLEE M M, et al. Microglial inflammatory signaling orchestrates the hypothalamic immune response to dietary excess and mediates obesity susceptibility[J]. Cell Metab, 2018, 27(6): 1356.
KIM J D, YOON N A, JIN S, et al. Microglial UCP2 mediates inflammation and obesity induced by high-fat feeding[J]. Cell Metab, 2019, 30(5): 952-962.e5.
CATERINA M J, SCHUMACHER M A, TOMINAGA M, et al. The capsaicin receptor: a heat-activated ion channel in the pain pathway[J]. Nature, 1997, 389(6653): 816-824.
MARRONE M C, MORABITO A, GIUSTIZIERI M, et al. TRPV1 channels are critical brain inflammation detectors and neuropathic pain biomarkers in mice[J]. Nat Commun, 2017, 8: 15292.
GIBSON H E, EDWARDS J G, PAGE R S, et al. TRPV1 channels mediate long-term depression at synapses on hippocampal interneurons[J]. Neuron, 2008, 57(5): 746-759.
MARINELLI S, MARZO V, BERRETTA N, et al. Presynaptic facilitation of glutamatergic synapses to dopaminergic neurons of the rat substantia nigra by endogenous stimulation of vanilloid receptors[J]. J Neurosci, 2003, 23(8): 3136-3144.
DOYLE M W, BAILEY T W, JIN Y H, et al. Vanilloid receptors presynaptically modulate cranial visceral afferent synaptic transmission in nucleus tractus solitarius[J]. J Neurosci, 2002, 22(18): 8222-8229.
KIM S R, KIM S U, OH U, et al. Transient receptor potential vanilloid subtype 1 mediates microglial cell death in vivo and in vitro via Ca2+-mediated mitochondrial damage and cytochrome c release[J]. J Immunol, 2006, 177(7): 4322-4329.
SAPPINGTON R M, CALKINS D J. Contribution of TRPV1 to microglia-derived IL-6 and NFkappaB translocation with elevated hydrostatic pressure[J]. Invest Ophthalmol Vis Sci, 2008, 49(7): 3004-3017.
SCHILLING T, EDER C. Importance of the non-selective cation channel TRPV1 for microglial reactive oxygen species generation[J]. J Neuroimmunol, 2009, 216(1/2): 118-121.
GAO W, SUN Y H, CAI M, et al. Copper sulfide nanoparticles as a photothermal switch for TRPV1 signaling to attenuate atherosclerosis[J]. Nat Commun, 2018, 9(1): 231.
WEI T J, WANG Y X, XU W R, et al. KCa3.1 deficiency attenuates neuroinflammation by regulating an astrocyte phenotype switch involving the PI3K/AKT/GSK3β pathway[J]. Neurobiol Dis, 2019, 132: 104588.
SHANNON P, MARKIEL A, OZIER O, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks[J]. Genome Res, 2003, 13(11): 2498-2504.
ZHOU Y Y, ZHOU B, PACHE L, et al. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets[J]. Nat Commun, 2019, 10(1): 1523.
FALK T, YUE X, ZHANG S L, et al. Vascular endothelial growth factor-B is neuroprotective in an in vivo rat model of Parkinson's disease[J]. Neurosci Lett, 2011, 496(1): 43-47.
KORDOWER J H, EMBORG M E, BLOCH J, et al. Neurodegeneration prevented by lentiviral vector delivery of GDNF in primate models of Parkinson's disease[J]. Science, 2000, 290(5492): 767-773.
ARENA E T, RUEDEN C T, HINER M C, et al. Quantitating the cell: turning images into numbers with ImageJ[J]. Wiley Interdiscip Rev Dev Biol, 2017, 6(2): 10.1002/wdev.260.
TRIEBL A, TRÖTZMÜLLER M, HARTLER J, et al. Lipidomics by ultrahigh performance liquid chromatography-high resolution mass spectrometry and its application to complex biological samples[J]. J Chromatogr B Analyt Technol Biomed Life Sci, 2017, 1053: 72-80.
TRACEY T J, STEYN F J, WOLVETANG E J, et al. Neuronal lipid metabolism: multiple pathways driving functional outcomes in health and disease[J]. Front Mol Neurosci, 2018, 11: 10.
MONNI M, CORAZZI L, MIGLIORATI G, et al. Respiratory state and phosphatidylserine import in brain mitochondria in vitro[J]. J Membrane Biol, 2000, 173(2): 97-105.
SHAHID R A, VIGNA S R, LAYNE A C, et al. Acinar cell production of leukotriene B4 contributes to development of neurogenic pancreatitis in mice[J]. Cell Mol Gastroenterol Hepatol, 2015, 1(1): 75-86.
MA L Q, ZHONG J, ZHAO Z G, et al. Activation of TRPV1 reduces vascular lipid accumulation and attenuates atherosclerosis[J]. Cardiovasc Res, 2011, 92(3): 504-513.
LI L, CHEN J, NI Y X, et al. TRPV1 activation prevents nonalcoholic fatty liver through UCP2 upregulation in mice[J]. Pflugers Arch - Eur J Physiol, 2012, 463(5): 727-732.
ZHAO J F, CHING L C, KOU Y R, et al. Activation of TRPV1 prevents OxLDL-induced lipid accumulation and TNF-α-induced inflammation in macrophages: role of liver X receptor Α[J]. Mediators Inflamm, 2013, 2013: 925171.
LEE E, JUNG D Y, KIM J H, et al. Transient receptor potential vanilloid type-1 channel regulates diet-induced obesity, insulin resistance, and leptin resistance[J]. FASEB J, 2015, 29(8): 3182-3192.
RAZAVI R, CHAN Y, AFIFIYAN F N, et al. TRPV1+ sensory neurons control beta cell stress and islet inflammation in autoimmune diabetes[J]. Cell, 2006, 127(6): 1123-1135.
FERNANDES E S, BRITO C X L, TEIXEIRA S A, et al. TRPV1 antagonism by capsazepine modulates innate immune response in mice infected with Plasmodium berghei ANKA[J]. Mediators Inflamm, 2014, 2014: 506450.
MANES T D, WANG V, POBER J S. Divergent TCR-initiated calcium signals govern recruitment versus activation of human alloreactive effector memory T cells by endothelial cells[J]. J Immunol, 2018, 201(11): 3167-3174.
HUANG W X, YU F, SANCHEZ R M, et al. TRPV1 promotes repetitive febrile seizures by pro-inflammatory cytokines in immature brain[J]. Brain Behav Immun, 2015, 48: 68-77.
YOSHIDA A, FURUBE E, MANNARI T, et al. TRPV1 is crucial for proinflammatory STAT3 signaling and thermoregulation-associated pathways in the brain during inflammation[J]. Sci Rep, 2016, 6: 26088.
CHEN Y, WILLCOCKSON H H, VALTSCHANOFF J G. Influence of the vanilloid receptor TRPV1 on the activation of spinal cord glia in mouse models of pain[J]. Exp Neurol, 2009, 220(2): 383-390.
HO K W, WARD N J, CALKINS D J. TRPV1: a stress response protein in the central nervous system[J]. Am J Neurodegener Dis, 2012, 1(1): 1-14.
KONG W L, PENG Y Y, PENG B W. Modulation of neuroinflammation: role and therapeutic potential of TRPV1 in the neuro-immune axis[J]. Brain Behav Immun, 2017, 64: 354-366.
LEONELLI M, MARTINS D O, BRITTO L R G. TRPV1 receptors are involved in protein nitration and Müller cell reaction in the acutely axotomized rat retina[J]. Exp Eye Res, 2010, 91(5): 755-768.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}