Analysis of transcriptome and chromatin accessibility changes during the differentiation of human embryonic stem cells into neural progenitor cells
LI Linying1, CAI Xiaodong2, TONG Ran2, YANG Chen2, WANG Zhiming2, HE Xiaoyu2, MA Ziyue2, ZHANG Feng,2, LI Lingjie,2, ZHOU Junmei,1
1.Department of Central Laboratory, Shanghai Children's Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai 200062, China
2.Department of Histoembryology, Genetics and Developmental Biology, Shanghai Jiao Tong University College of Basic Medical Sciences; Shanghai Key Laboratory of Reproductive Medicine; Key Laboratory of Cell Differentiation and Apoptosis of Chinese Ministry of Education, Shanghai 200025, China
目的·利用人胚胎干细胞(human embryonic stem cell,hESC)体外分化模型和高通量多组学测序技术研究hESC分化成神经前体细胞(neural progenitor cell,NPC)过程中转录组和染色质可及性的变化情况。方法·首先在体外利用拟胚体形成法诱导hESC分化成NPC,并收集这2个阶段的细胞;通过反转录-实时荧光定量PCR(reverse transcription-quantitative real-time PCR,RT-qPCR)和免疫荧光染色(immunofluorescence,IF)鉴定细胞表型。应用转录组测序(transcriptome sequencing,RNA-seq)检测并分析hESC和NPC的差异表达基因(differentially expressed gene,DEG)。应用染色质可及性测序(assay for transposase-accessible chromatin with high throughput sequencing,ATAC-seq)技术获取hESC和NPC的染色质可及性变化情况,并对差异的染色质开放区域进行基序富集分析以发现具有潜在调控作用的转录因子。最后对RNA-seq和ATAC-seq多组学数据进行联合分析,并构建蛋白质互作(protein-protein interaction,PPI)网络,寻找体外神经早期分化过程中的关键基因和调控通路。结果·RT-qPCR与IF均显示多能性标志物(NANOG、POU5F1)在hESC阶段表达量高而在NPC阶段表达量明显降低;同时神经早期分化标志物(PAX6、SOX1、NES)在hESC阶段基本不表达而在NPC阶段表达量显著升高。RNA-seq分析结果显示,与hESC阶段相比,NPC阶段中有5 597个基因的表达水平呈现上调,而3 654个基因的表达水平下降,基因功能富集分析显示NPC阶段上调的基因富集至神经发育相关的功能。ATAC-seq分析结果显示,共27 491个基因组区域在hESC向NPC分化过程中染色质可及性发生了显著改变,其中有12 381个区域染色质可及性增强,15 110个区域染色质可及性减弱;基序富集分析揭示DLX1、LHX2等转录因子基因可能在hESC向NPC分化过程中发挥重要作用。RNA-seq和ATAC-seq的多组学数据联合分析结果显示,在NPC阶段高表达的重叠基因主要富集在轴突导向、前脑发育、神经元迁移等。神经分化后CTNND2、LHX2基因表达水平升高,且相关基因区域染色质可及性也增加。PPI网络分析发现,PRKACA、CDH2、ERBB4等是下游候选基因。结论·利用hESC体外分化模型结合高通量多组学测序技术可用于揭示hESC向NPC分化过程中的转录组及染色质可及性的变化规律;该过程中轴突导向、前脑发育、神经元迁移等通路的相关基因表达水平升高,染色质可及性增强。
关键词:人胚胎干细胞
;
神经前体细胞
;
染色质可及性测序
;
转录组测序
Abstract
Objective ·To investigate the changes in transcriptome and chromatin accessibility during the differentiation of human embryonic stem cells (hESCs) into neural progenitor cells (NPCs) using in vitro differentiation models and high-throughput multi-omics sequencing technologies. Methods ·hESCs were first induced to differentiate into NPCs in vitro using the embryoid body formation method, and cells at both stages were collected. The cell phenotypes were identified by reverse transcription-quantitative real-time PCR (RT-qPCR) and immunofluorescence (IF) staining. Transcriptome sequencing (RNA-seq) was conducted to detect and analyze the differentially expressed genes (DEGs) between hESCs and NPCs. The assay for transposase-accessible chromatin with high-throughput sequencing (ATAC-seq) was employed to assess chromatin accessibility changes between hESCs and NPCs. Motif enrichment analysis was performed on differentially accessible chromatin regions to discover potential regulatory transcription factors. Finally, an integrated analysis of RNA-seq and ATAC-seq data and the protein-protein interaction (PPI) network were performed to identify key genes and regulatory pathways involved in the early stages of neural differentiation in vitro. Results ·Both RT-qPCR and IF results indicated that the expression levels of pluripotency markers (NANOG and POU5F1) were high at the hESC stage but significantly decreased at the NPC stage, while early neural differentiation markers (PAX6, SOX1, and NES) were minimally expressed at the hESC stage but markedly upregulated at the NPC stage. RNA-seq analysis revealed that compared to the hESC stage, there were 5 597 genes upregulated and 3 654 genes downregulated at the NPC stage. Gene function enrichment analysis showed that the upregulated genes at the NPC stage were enriched in the functions related to neural development. ATAC-seq analysis demonstrated a total of 27 491 genomic regions had significant changes in chromatin accessibility during the differentiation from hESC to NPC, with 12 381 regions showing increased accessibility and 15 110 regions showing decreased accessibility. Motif enrichment analysis revealed that transcription factor genes such as DLX1 and LHX2 might play an important role in the differentiation process from hESCs into NPCs. Integrated analysis of RNA-seq and ATAC-seq data revealed that overlapping genes with high expression at the NPC stage were mainly enriched in axon guidance, forebrain development, and neuron migration. After neural differentiation, the expression levels of CTNND2 and LHX2 genes increased, and the chromatin accessibility of related genomic regions also increased. PPI network analysis indentified candidate downstream genes including PRKACA, CDH2, and ERBB4. Conclusion ·The in vitro differentiation model of hESCs combined with high-throughput multi-omics sequencing technologies can be used to depict the changes in transcriptome and chromatin accessibility during the differentiation of hESCs into NPCs. In this process, the expression levels of genes related to axon guidance, forebrain development, and neuronal migration pathways increase and related chromatin accessibility is enhanced.
LI Linying, CAI Xiaodong, TONG Ran, YANG Chen, WANG Zhiming, HE Xiaoyu, MA Ziyue, ZHANG Feng, LI Lingjie, ZHOU Junmei. Analysis of transcriptome and chromatin accessibility changes during the differentiation of human embryonic stem cells into neural progenitor cells. Journal of Shanghai Jiao Tong University (Medical Science)[J], 2025, 45(4): 387-403 doi:10.3969/j.issn.1674-8115.2025.04.001
细胞命运决定直接影响着不同细胞类型的分化和特化。通过正确的细胞命运决定过程,原始细胞能够选择合适的分化路径,并最终形成正确的细胞类型。在发育中的大脑皮层,神经前体细胞(neural progenitor cell,NPC)通过自我更新及增殖,具备不同的谱系特点,并受到精确调控,最终分化为神经元、少突胶质细胞和星形胶质细胞三大神经细胞类型[1-3]。研究发现,神经细胞亚型的异常在不同的神经精神疾病中存在。例如,利用自闭症谱系障碍(autism spectrum disorders,ASD)患者来源的多能干细胞(pluripotent stem cell,PSC)分化出NPC,可检测到谷氨酸能神经细胞比例下降,而γ-氨基丁酸能神经细胞比例增加,提示兴奋性和抑制性的NPC比例失衡[4];这一发现与ASD的重要病因假说,即神经元回路中兴奋/抑制平衡(balance of excitation and inhibition,E/I balance)的失调[5-7]相吻合。因此,探索神经细胞如何分化产生丰富的细胞类型,最终不同细胞类型如何以高度准确的时空规律组装成复杂神经环路,都具有极其重要的意义。
染色质可及性(chromatin accessibility)是指细胞核内大分子能够与染色质DNA物理接触的程度[16];在转录或复制合成过程中,核小体从DNA上解离,使DNA暴露以便能够被转录调控因子结合,启动转录并调控基因的表达。通过检测染色质可及性,可以直接反映出调控因子与开放染色质结合的状态,该状态与基因的表达情况密切相关。目前研究染色质可及性的方法主要是将酶切法或物理化学方法与染色质可及性测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)结合,以识别染色质开放区域或核小体占据的区域[17-18]。近年来,有大量研究[19-20]利用检测染色质可及性的技术研究了神经发育过程中表观基因组的动态变化。
所有的RNA-seq原始数据均以FASTQ格式下机。使用Cutadapt软件(1.9.1版本)去除接头序列[29],用Trimmomatic软件(0.3.2版本)[30]删去低质量读段,保留高质量的测序读段用于后续的分析。使用FastQC软件(0.11.9版本)[31]对下机数据进行质量控制。下载参考基因组和基因注释文件,使用HISAT2软件(2.0.5版本)[32]构建参考基因组的索引,并与参考基因组hg19进行比对;根据RefSeq转录本注释确定转录水平,并使用featureCounts程序(1.5.0-p3版本)[33]对能够比对到基因的读段数量进行统计,使用FPKM(fragments per kilobase of exon model per million mapped fragments)对表达量进行标准化,以使不同基因和样本之间的表达水平具有可比性;使用DESeq软件(1.39.0版本)[34]对不同组间的基因表达进行差异分析,差异表达基因(differentially expressed gene,DEG)被定义为至少一个重复的FPKM>1,且差异倍数(fold change,FC)对数的绝对值()>1,P<0.05。使用ClusterProfiler软件(4.10.1版本)对DEG进行功能富集分析[35]。本研究中所有RNA-seq原始测序数据均已上传至国家生物信息中心(https://www.cncb.ac.cn/),GSA-Human编号为HRA009071。
1.2.7 ATAC-seq数据分析
首先使用Trimmomatic软件(0.3.2版本)[30]对原始测序数据进行质量控制,包括去除低质量的碱基、修剪接头序列和过滤掉低质量的读段;过滤掉的数据用FastQC软件(0.11.9版本)[31]进行质量控制;之后使用工具Bowtie2软件(2.5.2版本)将质控过的测序数据与参考基因组hg19进行比对[36],生成BAM文件,该文件包含了每个读段的比对位置信息。使用MACS2软件(2.1.4版本)识别基因组上染色质开放区域显著富集的位点或峰[37],并用MEME软件(5.5.7版本)[38]对这些峰信号进行注释;利用DiffBind软件(2.10版本)[39]对不同样本的ATAC-seq数据进行差异分析,以确定差异的染色质开放区域。利用基序(motif)的序列分析工具(MEME中的AME工具)进行相关富集分析,比对的基序数据库为JASPAR2022 CORE non-redundant v2,比对基因组为hg19。利用GREAT(Genomic Regions Enrichment of Annotations Tool)在线网站(http://great.stanford.edu/public/html/)对差异染色质开放区域进行功能富集分析,以了解这些区域所涉及的生物学过程、通路和功能。常用的富集分析工具包括GO(Gene Ontology)、KEGG(Kyoto Encyclopedia of Genes and Genomes)和GREAT等。本研究中所有ATAC-seq原始测序数据均已上传至国家生物信息中心(https://www.cncb.ac.cn/),GSA-Human编号为HRA009070。
1.2.8 构建蛋白质互作网络
将基因集导入交互基因检索工具(Search Tool for the Retrieval of Interacting Genes,STRING;http://string-db.org),选择物种为Homo sapiens,设置置信度阈值大于0.7,从而得到该基因集的蛋白质–蛋白质相互作用(protein-protein interaction,PPI)网络;之后通过Cytoscape软件(3.7.2版本)进行可视化和网络分析,将筛选出的度(degree)≥10的节点作为PPI网络的重要基因。
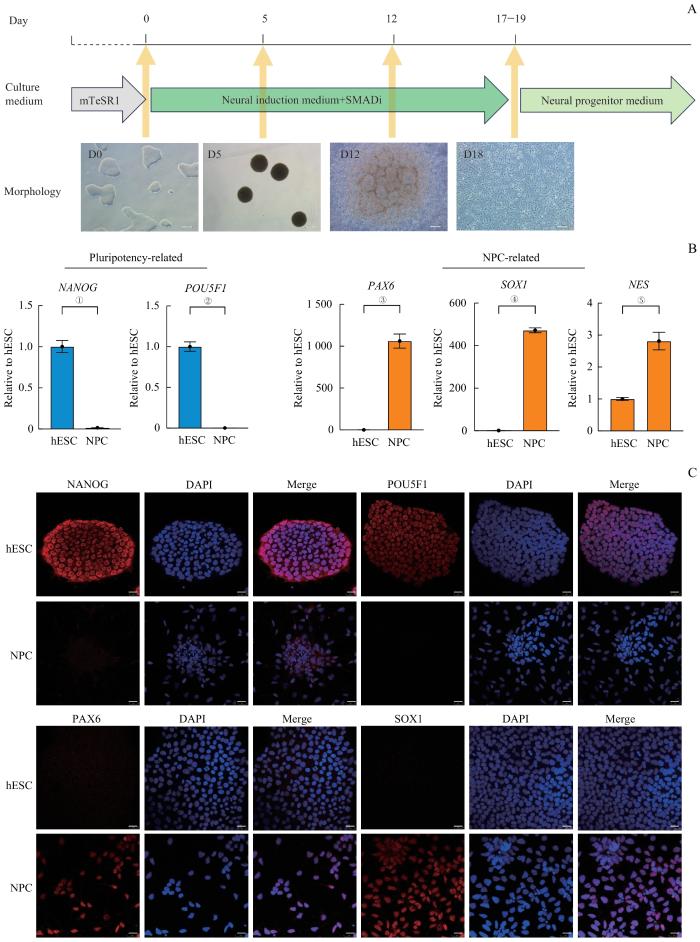
Note:A. Schematic diagram of the NPC culture system and morphology of hESC-induced differentiation into NPCs. SMADi—SMAD inhibition. Scale bar=200 μm. B. The gene expression of typical genes related to pluripotency and NPCs during the induction process detected by RT-qPCR. ①P=0.002, ②P=0.001, ③P=0.002, ④P<0.001, ⑤P=0.006. NPCs were the cells induced for 27 d. C. Expression of hESC markers (NANOG and POU5F1) and NPC markers (PAX6 and SOX1), detected by immunofluorescence staining. NPCs were the cells induced for 27 d. Scale bar=25 μm.
Fig 1
Dual SMAD inhibition strategy for hESC differentiation into NPC in vitro
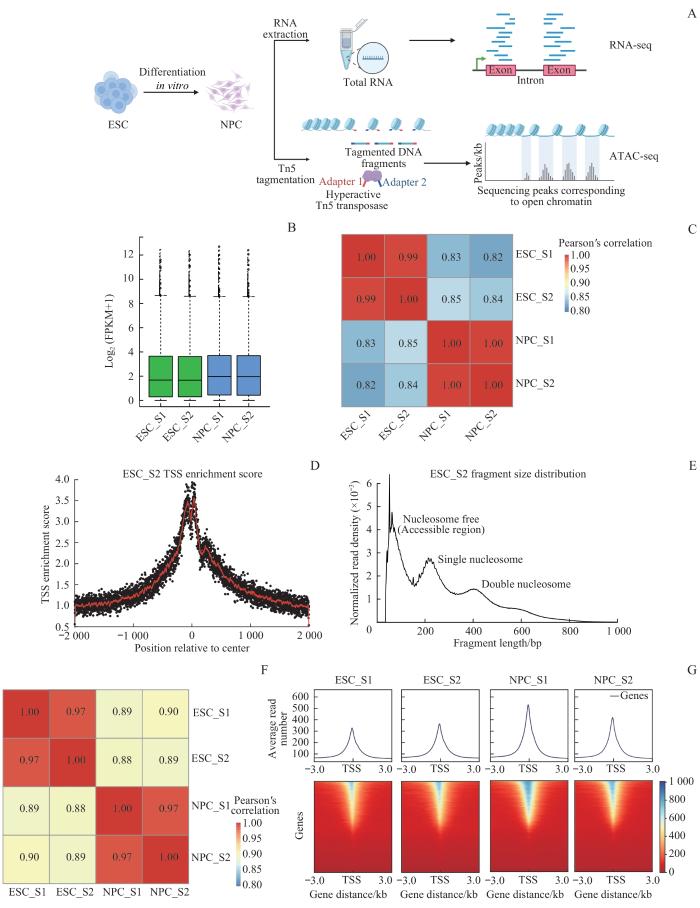
Note: A. Experimental flow chart. Cells were collected at the ESC stage and NPC stage; nuclei were extracted for standard ATAC-seq library preparation, while total RNA was extracted for RNA-seq library preparation. B. Box plot of gene expression levels. C. Pearson correlation analysis was performed on the expression levels between different samples. D. Transcription start site (TSS) enrichment scores of ATAC-seq data. E. Fragment size distribution of ATAC-seq data. F. Pearson correlation analysis of ATAC-seq data between different samples. G. Gene TSS enrichment heat map showing the reads from ATAC-seq data in the TSS±3.0 kb. The blue-to-red scale represents the transition from high to low in the reads in the specified area.
Fig 2
Quality control analysis of RNA-seq and ATAC-seq datasets obtained from the process of early neural differentiation in vitro
Tab 2
表2
表2在神经诱导过程中RNA-seq文库所获得的原始读段数和比对读段数
Tab 2 Number of raw reads and mapped reads obtained from RNA-seq libraries during neural induction
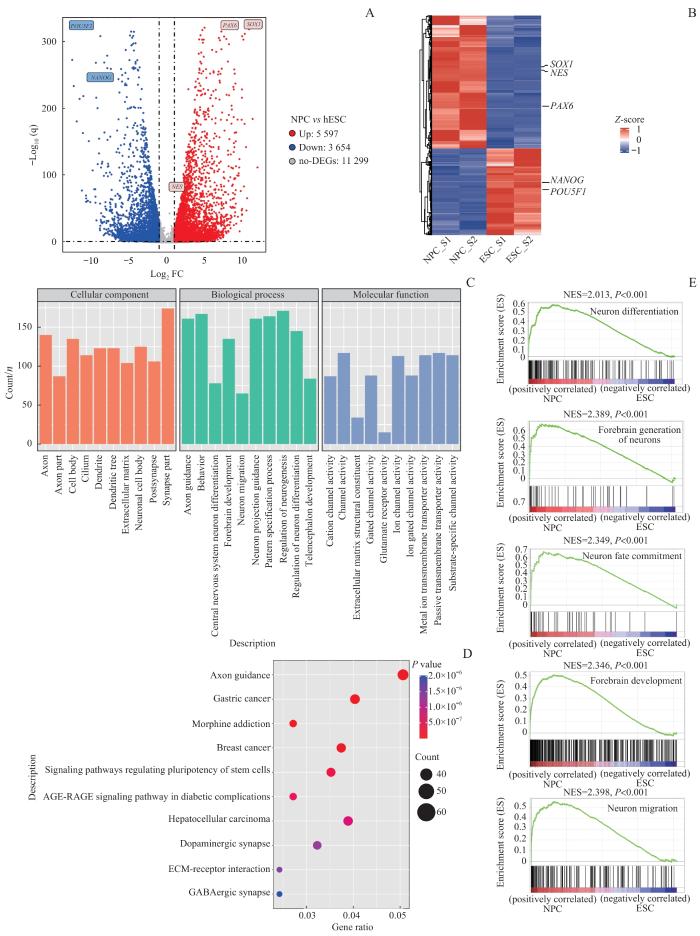
Note: A. Volcano plot shows the distribution of DEGs between the two distinct developmental stages (ESC and NPC). DEGs were defined as |log2FC|>1 and P<0.05. B. Gene expression levels were clustered across all samples. Different colors indicate the value of each gene expression level normalized with Z-scores. C. GO analysis was performed on the DEGs upregulated at the NPC stage. D. KEGG analysis of the DEGs upregulated at NPC stage. E. GSEA analysis identifies pathways upregulated during NPC differentiation.
Fig 3
Transcriptome analysis of NPC differentiation
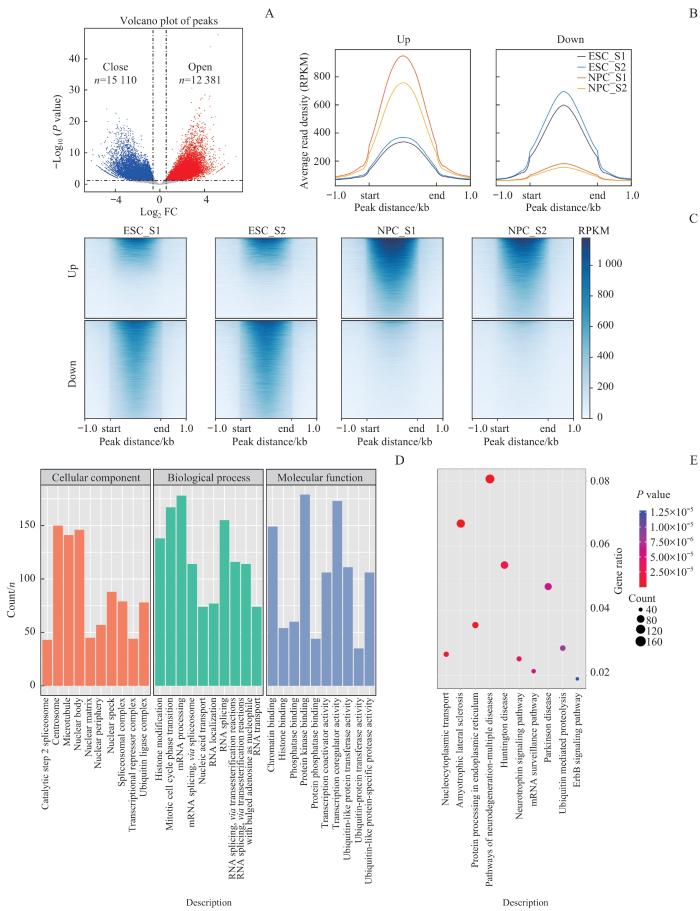
Note: A. Volcano plot shows the peak signals of differentially open chromatin obtained from the ATAC-seq dataset. Differential open areas were defined as |log2FC| >0.585 and P<0.05. B. The differential region enrichment plot shows the average peak signal between different samples in the differentially upregulated region (left) and the differentially downregulated region (right) at the NPC stage. C. Differential region enrichment heatmap reveals peak signals within genomic region. D. GO analysis of genes associated with differentially open chromatin regions at the NPC stage. E. KEGG analysis of genes associated with differentially open chromatin regions at the NPC stage.
Fig 4
Chromatin accessibility analysis during NPC differentiation process
Tab 4
表4
表4对在NPC差异上调的开放染色质信号进行基序富集分析
Tab 4 Motif enrichment analysis of open chromatin signals that were differentially upregulated at the NPC stage
Motif
E value
Motif ID
Motif
E value
Motif ID
Note:DLX1——distal-less homeobox 1; SHOX——SHOX homeobox; MSX1——MSH homeobox 1; NKX6.1——NK6 homeobox 1; LHX2—LIM homeobox 2. The relative size of each position letter in motif indicates their frequency in the sequence.
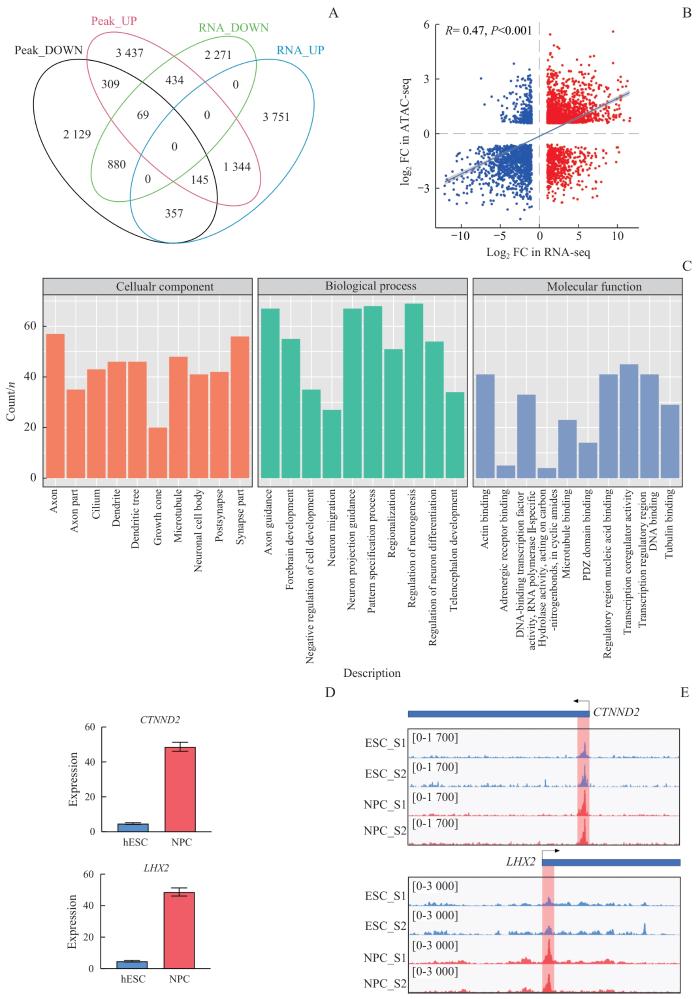
Note: A. Venn diagram showing the overlap between DEGs in the NPC cell population and related genes predicted from differentially open chromatin regions. In the ATAC-seq dataset, genes were annotated to regions with ±1 kb of the promoter. B. Scatter plot showing the correlation between the expression levels of the overlapping genes (A) obtained in RNA-seq and the associated peaks obtained from ATAC-seq. C. GO analysis of 1 344 upregulated genes. D. Expression levels of CTNND2 and LHX2 in RNA-seq during NPC differentiation. E. The Integrative Genomics Viewer showing ATAC-seq signals at the transcription start sites of CTNND2 and LHX2 loci during NPC differentiation.
Fig 5
Integration of RNA-seq and ATAC-seq data to analyze important genes involved in NPC differentiation
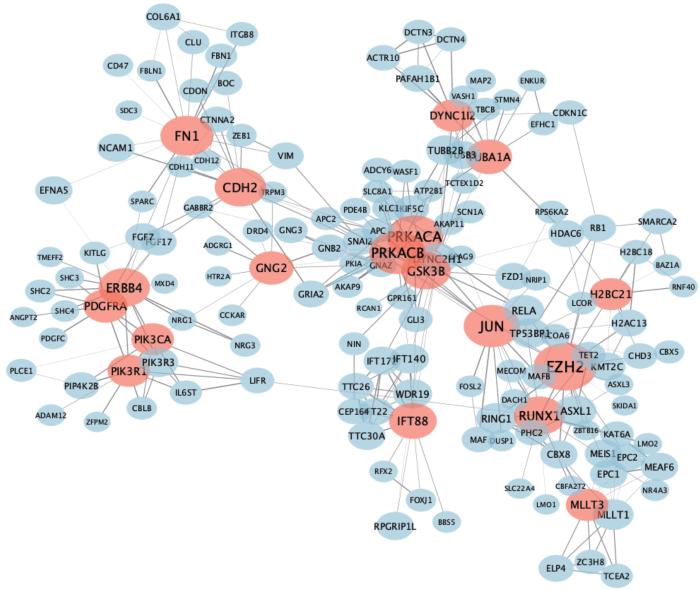
Note: The thickness of the edge in the network diagram represents the interaction strength between the two proteins, and the size of the node represents the degree. The red circles represent the protein with degree≥10, and the others represent proteins that interact with the red proteins.
Fig 6
Candidate hub downstream genes detected by PPI network analysis
Tab 5
表5
表5PPI网络中度≥10的下游候选中心基因
Tab 5 Candidate downstream hub genes with the degrees≥10 in the PPI network
The study was designed by LI Lingjie, ZHOU Junmei and ZHANG Feng. The experiments were performed, and the data were collected and analyzed by LI Linying. CAI Xiaodong and TONG Ran contributed to the experiment operation. YANG Chen, WANG Zhiming, HE Xiaoyu, and MA Ziyue contributed to the data analysis. The manuscript was drafted and revised by all the authors. All authors have read the final version of paper and consented to submission.
利益冲突声明
所有作者声明不存在利益冲突。
COMPETING INTERESTS
All authors declare no relevant conflict of interests.
NOCTOR S C, FLINT A C, WEISSMAN T A, et al. Neurons derived from radial glial cells establish radial units in neocortex[J]. Nature, 2001, 409(6821): 714-720.
KRIEGSTEIN A, ALVAREZ-BUYLLA A. The glial nature of embryonic and adult neural stem cells[J]. Annu Rev Neurosci, 2009, 32: 149-184.
MARCHETTO M C, BELINSON H, TIAN Y, et al. Altered proliferation and networks in neural cells derived from idiopathic autistic individuals[J]. Mol Psychiatry, 2017, 22(6): 820-835.
HOWELL B W, SMITH K M. Synaptic structural protein dysfunction leads to altered excitation inhibition ratios in models of autism spectrum disorder[J]. Pharmacol Res, 2019, 139: 207-214.
BOZZI Y, PROVENZANO G, CASAROSA S. Neurobiological bases of autism-epilepsy comorbidity: a focus on excitation/inhibition imbalance[J]. Eur J Neurosci, 2018, 47(6): 534-548.
OLIVEIRA B, MITJANS M, NITSCHE M A, et al. Excitation-inhibition dysbalance as predictor of autistic phenotypes[J]. J Psychiatr Res, 2018, 104: 96-99.
DAI S K, LIU P P, LI X, et al. Dynamic profiling and functional interpretation of histone lysine crotonylation and lactylation during neural development[J]. Development, 2022, 149(14): dev200049.
SUN T Y, XU Y Y, XIANG Y, et al. Crosstalk between RNA m6A and DNA methylation regulates transposable element chromatin activation and cell fate in human pluripotent stem cells[J]. Nat Genet, 2023, 55(8): 1324-1335.
WANG C F, YANG J W, ZHUANG Z H, et al. Activity-dependent feedback regulation of thalamocortical axon development by Lhx2 in cortical layer 4 neurons[J]. Cereb Cortex, 2023, 33(5): 1693-1707.
BUENROSTRO J D, GIRESI P G, ZABA L C, et al. Transposition of native chromatin for fast and sensitive epigenomic profiling of open chromatin, DNA-binding proteins and nucleosome position[J]. Nat Methods, 2013, 10(12): 1213-1218.
MARKENSCOFF-PAPADIMITRIOU E, WHALEN S, PRZYTYCKI P, et al. A chromatin accessibility atlas of the developing human telencephalon[J]. Cell, 2020, 182(3): 754-769.e18.
TAKAHASHI K, TANABE K, OHNUKI M, et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors[J]. Cell, 2007, 131(5): 861-872.
YU J Y, VODYANIK M A, SMUGA-OTTO K, et al. Induced pluripotent stem cell lines derived from human somatic cells[J]. Science, 2007, 318(5858): 1917-1920.
CHAMBERS S M, FASANO C A, PAPAPETROU E P, et al. Highly efficient neural conversion of human ES and iPS cells by dual inhibition of SMAD signaling[J]. Nat Biotechnol, 2009, 27(3): 275-280.
KAWASAKI H, MIZUSEKI K, NISHIKAWA S, et al. Induction of midbrain dopaminergic neurons from ES cells by stromal cell-derived inducing activity[J]. Neuron, 2000, 28(1): 31-40.
LEE H, SHAMY G A, ELKABETZ Y, et al. Directed differentiation and transplantation of human embryonic stem cell-derived motoneurons[J]. Stem Cells, 2007, 25(8): 1931-1939.
ZHANG S C, WERNIG M, DUNCAN I D, et al. In vitro differentiation of transplantable neural precursors from human embryonic stem cells[J]. Nat Biotechnol, 2001, 19(12): 1129-1133.
TCHIEU J, ZIMMER B, FATTAHI F, et al. A modular platform for differentiation of human PSCs into all major ectodermal lineages[J]. Cell Stem Cell, 2017, 21(3): 399-410.e7.
LIAO Y, SMYTH G K, SHI W. featureCounts: an efficient general purpose program for assigning sequence reads to genomic features[J]. Bioinformatics, 2014, 30(7): 923-930.
BAILEY T L, BODEN M, BUSKE F A, et al. MEME SUITE: tools for motif discovery and searching[J]. Nucleic Acids Res, 2009, 37(Web Server issue): W202-W208.
ROSS-INNES C S, STARK R, TESCHENDORFF A E, et al. Differential oestrogen receptor binding is associated with clinical outcome in breast cancer[J]. Nature, 2012, 481(7381): 389-393.
WATANABE K, UENO M, KAMIYA D, et al. A ROCK inhibitor permits survival of dissociated human embryonic stem cells[J]. Nat Biotechnol, 2007, 25(6): 681-686.
ITSKOVITZ-ELDOR J, SCHULDINER M, KARSENTI D, et al. Differentiation of human embryonic stem cells into embryoid bodies compromising the three embryonic germ layers[J]. Mol Med, 2000, 6(2): 88-95.
SZKLARCZYK D, KIRSCH R, KOUTROULI M, et al. The STRING database in 2023: protein-protein association networks and functional enrichment analyses for any sequenced genome of interest[J]. Nucleic Acids Res, 2023, 51(D1): D638-D646.
JIA E T, PAN M, LIU Z Y, et al. Transcriptomic profiling of differentially expressed genes and related pathways in different brain regions in Parkinson's disease[J]. Neurosci Lett, 2020, 732: 135074.
ARAYA C, HÄKKINEN H M, CARCAMO L, et al. Cdh2 coordinates Myosin-Ⅱ dependent internalisation of the zebrafish neural plate[J]. Sci Rep, 2019, 9(1): 1835.
BARETTINO C, BALLESTEROS-GONZALEZ Á, AYLÓN A, et al. Developmental disruption of Erbb4 in Pet1+ neurons impairs serotonergic sub-system connectivity and memory formation[J]. Front Cell Dev Biol, 2021, 9: 770458.
TUNCAY I O, PARMALEE N L, KHALIL R, et al. Analysis of recent shared ancestry in a familial cohort identifies coding and noncoding autism spectrum disorder variants[J]. NPJ Genom Med, 2022, 7(1): 13.
ASADOLLAHI R, ONEDA B, JOSET P, et al. The clinical significance of small copy number variants in neurodevelopmental disorders[J]. J Med Genet, 2014, 51(10): 677-688.
CHEN Z N, LI X J, CUI X Z, et al. Association of CTNND2 gene polymorphism with schizophrenia: two-sample case-control study in Chinese Han population[J]. Int J Psychiatry Med, 2023, 58(5): 433-448.
WANG L Y, XU M, WANG Y, et al. Melatonin improves synapse development by PI3K/Akt signaling in a mouse model of autism spectrum disorder[J]. Neural Regen Res, 2024, 19(7): 1618-1624.
VAZ R, EDWARDS S, DUEÑAS-REY A, et al. Loss of ctnnd2b affects neuronal differentiation and behavior in zebrafish[J]. Front Neurosci, 2023, 17: 1205653.
WANG Y N, KHANDELWAL N, LIU S Q, et al. KDM6B cooperates with Tau and regulates synaptic plasticity and cognition via inducing VGLUT1/2[J]. Mol Psychiatry, 2022, 27(12): 5213-5226.
LI W, SHEN W C, ZHANG B, et al. Long non-coding RNA LncKdm2b regulates cortical neuronal differentiation by cis-activating Kdm2b[J]. Protein Cell, 2020, 11(3): 161-186.
CHEN D H, MCMANUS C E, RADMANESH B, et al. Temporal inhibition of chromatin looping and enhancer accessibility during neuronal remodeling[J]. Nat Commun, 2021, 12(1): 6366.
RAHMAN S, DONG P F, APONTES P, et al. Lineage specific 3D genome structure in the adult human brain and neurodevelopmental changes in the chromatin interactome[J]. Nucleic Acids Res, 2023, 51(20): 11142-11161.
CORCES M R, SHCHERBINA A, KUNDU S, et al. Single-cell epigenomic analyses implicate candidate causal variants at inherited risk loci for Alzheimer's and Parkinson's diseases[J]. Nat Genet, 2020, 52(11): 1158-1168.
PAGNI S, MILLS J D, FRANKISH A, et al. Non-coding regulatory elements: potential roles in disease and the case of epilepsy[J]. Neuropathol Appl Neurobiol, 2022, 48(3): e12775.
... 染色质可及性(chromatin accessibility)是指细胞核内大分子能够与染色质DNA物理接触的程度[16];在转录或复制合成过程中,核小体从DNA上解离,使DNA暴露以便能够被转录调控因子结合,启动转录并调控基因的表达.通过检测染色质可及性,可以直接反映出调控因子与开放染色质结合的状态,该状态与基因的表达情况密切相关.目前研究染色质可及性的方法主要是将酶切法或物理化学方法与染色质可及性测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)结合,以识别染色质开放区域或核小体占据的区域[17-18].近年来,有大量研究[19-20]利用检测染色质可及性的技术研究了神经发育过程中表观基因组的动态变化. ...
1
... 染色质可及性(chromatin accessibility)是指细胞核内大分子能够与染色质DNA物理接触的程度[16];在转录或复制合成过程中,核小体从DNA上解离,使DNA暴露以便能够被转录调控因子结合,启动转录并调控基因的表达.通过检测染色质可及性,可以直接反映出调控因子与开放染色质结合的状态,该状态与基因的表达情况密切相关.目前研究染色质可及性的方法主要是将酶切法或物理化学方法与染色质可及性测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)结合,以识别染色质开放区域或核小体占据的区域[17-18].近年来,有大量研究[19-20]利用检测染色质可及性的技术研究了神经发育过程中表观基因组的动态变化. ...
2
... 染色质可及性(chromatin accessibility)是指细胞核内大分子能够与染色质DNA物理接触的程度[16];在转录或复制合成过程中,核小体从DNA上解离,使DNA暴露以便能够被转录调控因子结合,启动转录并调控基因的表达.通过检测染色质可及性,可以直接反映出调控因子与开放染色质结合的状态,该状态与基因的表达情况密切相关.目前研究染色质可及性的方法主要是将酶切法或物理化学方法与染色质可及性测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)结合,以识别染色质开放区域或核小体占据的区域[17-18].近年来,有大量研究[19-20]利用检测染色质可及性的技术研究了神经发育过程中表观基因组的动态变化. ...
... 染色质可及性(chromatin accessibility)是指细胞核内大分子能够与染色质DNA物理接触的程度[16];在转录或复制合成过程中,核小体从DNA上解离,使DNA暴露以便能够被转录调控因子结合,启动转录并调控基因的表达.通过检测染色质可及性,可以直接反映出调控因子与开放染色质结合的状态,该状态与基因的表达情况密切相关.目前研究染色质可及性的方法主要是将酶切法或物理化学方法与染色质可及性测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)结合,以识别染色质开放区域或核小体占据的区域[17-18].近年来,有大量研究[19-20]利用检测染色质可及性的技术研究了神经发育过程中表观基因组的动态变化. ...
1
... 染色质可及性(chromatin accessibility)是指细胞核内大分子能够与染色质DNA物理接触的程度[16];在转录或复制合成过程中,核小体从DNA上解离,使DNA暴露以便能够被转录调控因子结合,启动转录并调控基因的表达.通过检测染色质可及性,可以直接反映出调控因子与开放染色质结合的状态,该状态与基因的表达情况密切相关.目前研究染色质可及性的方法主要是将酶切法或物理化学方法与染色质可及性测序(assay for transposase-accessible chromatin with high-throughput sequencing,ATAC-seq)结合,以识别染色质开放区域或核小体占据的区域[17-18].近年来,有大量研究[19-20]利用检测染色质可及性的技术研究了神经发育过程中表观基因组的动态变化. ...
... 所有的RNA-seq原始数据均以FASTQ格式下机.使用Cutadapt软件(1.9.1版本)去除接头序列[29],用Trimmomatic软件(0.3.2版本)[30]删去低质量读段,保留高质量的测序读段用于后续的分析.使用FastQC软件(0.11.9版本)[31]对下机数据进行质量控制.下载参考基因组和基因注释文件,使用HISAT2软件(2.0.5版本)[32]构建参考基因组的索引,并与参考基因组hg19进行比对;根据RefSeq转录本注释确定转录水平,并使用featureCounts程序(1.5.0-p3版本)[33]对能够比对到基因的读段数量进行统计,使用FPKM(fragments per kilobase of exon model per million mapped fragments)对表达量进行标准化,以使不同基因和样本之间的表达水平具有可比性;使用DESeq软件(1.39.0版本)[34]对不同组间的基因表达进行差异分析,差异表达基因(differentially expressed gene,DEG)被定义为至少一个重复的FPKM>1,且差异倍数(fold change,FC)对数的绝对值()>1,P<0.05.使用ClusterProfiler软件(4.10.1版本)对DEG进行功能富集分析[35].本研究中所有RNA-seq原始测序数据均已上传至国家生物信息中心(https://www.cncb.ac.cn/),GSA-Human编号为HRA009071. ...
2
... 所有的RNA-seq原始数据均以FASTQ格式下机.使用Cutadapt软件(1.9.1版本)去除接头序列[29],用Trimmomatic软件(0.3.2版本)[30]删去低质量读段,保留高质量的测序读段用于后续的分析.使用FastQC软件(0.11.9版本)[31]对下机数据进行质量控制.下载参考基因组和基因注释文件,使用HISAT2软件(2.0.5版本)[32]构建参考基因组的索引,并与参考基因组hg19进行比对;根据RefSeq转录本注释确定转录水平,并使用featureCounts程序(1.5.0-p3版本)[33]对能够比对到基因的读段数量进行统计,使用FPKM(fragments per kilobase of exon model per million mapped fragments)对表达量进行标准化,以使不同基因和样本之间的表达水平具有可比性;使用DESeq软件(1.39.0版本)[34]对不同组间的基因表达进行差异分析,差异表达基因(differentially expressed gene,DEG)被定义为至少一个重复的FPKM>1,且差异倍数(fold change,FC)对数的绝对值()>1,P<0.05.使用ClusterProfiler软件(4.10.1版本)对DEG进行功能富集分析[35].本研究中所有RNA-seq原始测序数据均已上传至国家生物信息中心(https://www.cncb.ac.cn/),GSA-Human编号为HRA009071. ...
... 首先使用Trimmomatic软件(0.3.2版本)[30]对原始测序数据进行质量控制,包括去除低质量的碱基、修剪接头序列和过滤掉低质量的读段;过滤掉的数据用FastQC软件(0.11.9版本)[31]进行质量控制;之后使用工具Bowtie2软件(2.5.2版本)将质控过的测序数据与参考基因组hg19进行比对[36],生成BAM文件,该文件包含了每个读段的比对位置信息.使用MACS2软件(2.1.4版本)识别基因组上染色质开放区域显著富集的位点或峰[37],并用MEME软件(5.5.7版本)[38]对这些峰信号进行注释;利用DiffBind软件(2.10版本)[39]对不同样本的ATAC-seq数据进行差异分析,以确定差异的染色质开放区域.利用基序(motif)的序列分析工具(MEME中的AME工具)进行相关富集分析,比对的基序数据库为JASPAR2022 CORE non-redundant v2,比对基因组为hg19.利用GREAT(Genomic Regions Enrichment of Annotations Tool)在线网站(http://great.stanford.edu/public/html/)对差异染色质开放区域进行功能富集分析,以了解这些区域所涉及的生物学过程、通路和功能.常用的富集分析工具包括GO(Gene Ontology)、KEGG(Kyoto Encyclopedia of Genes and Genomes)和GREAT等.本研究中所有ATAC-seq原始测序数据均已上传至国家生物信息中心(https://www.cncb.ac.cn/),GSA-Human编号为HRA009070. ...
2
... 所有的RNA-seq原始数据均以FASTQ格式下机.使用Cutadapt软件(1.9.1版本)去除接头序列[29],用Trimmomatic软件(0.3.2版本)[30]删去低质量读段,保留高质量的测序读段用于后续的分析.使用FastQC软件(0.11.9版本)[31]对下机数据进行质量控制.下载参考基因组和基因注释文件,使用HISAT2软件(2.0.5版本)[32]构建参考基因组的索引,并与参考基因组hg19进行比对;根据RefSeq转录本注释确定转录水平,并使用featureCounts程序(1.5.0-p3版本)[33]对能够比对到基因的读段数量进行统计,使用FPKM(fragments per kilobase of exon model per million mapped fragments)对表达量进行标准化,以使不同基因和样本之间的表达水平具有可比性;使用DESeq软件(1.39.0版本)[34]对不同组间的基因表达进行差异分析,差异表达基因(differentially expressed gene,DEG)被定义为至少一个重复的FPKM>1,且差异倍数(fold change,FC)对数的绝对值()>1,P<0.05.使用ClusterProfiler软件(4.10.1版本)对DEG进行功能富集分析[35].本研究中所有RNA-seq原始测序数据均已上传至国家生物信息中心(https://www.cncb.ac.cn/),GSA-Human编号为HRA009071. ...
... 首先使用Trimmomatic软件(0.3.2版本)[30]对原始测序数据进行质量控制,包括去除低质量的碱基、修剪接头序列和过滤掉低质量的读段;过滤掉的数据用FastQC软件(0.11.9版本)[31]进行质量控制;之后使用工具Bowtie2软件(2.5.2版本)将质控过的测序数据与参考基因组hg19进行比对[36],生成BAM文件,该文件包含了每个读段的比对位置信息.使用MACS2软件(2.1.4版本)识别基因组上染色质开放区域显著富集的位点或峰[37],并用MEME软件(5.5.7版本)[38]对这些峰信号进行注释;利用DiffBind软件(2.10版本)[39]对不同样本的ATAC-seq数据进行差异分析,以确定差异的染色质开放区域.利用基序(motif)的序列分析工具(MEME中的AME工具)进行相关富集分析,比对的基序数据库为JASPAR2022 CORE non-redundant v2,比对基因组为hg19.利用GREAT(Genomic Regions Enrichment of Annotations Tool)在线网站(http://great.stanford.edu/public/html/)对差异染色质开放区域进行功能富集分析,以了解这些区域所涉及的生物学过程、通路和功能.常用的富集分析工具包括GO(Gene Ontology)、KEGG(Kyoto Encyclopedia of Genes and Genomes)和GREAT等.本研究中所有ATAC-seq原始测序数据均已上传至国家生物信息中心(https://www.cncb.ac.cn/),GSA-Human编号为HRA009070. ...
1
... 所有的RNA-seq原始数据均以FASTQ格式下机.使用Cutadapt软件(1.9.1版本)去除接头序列[29],用Trimmomatic软件(0.3.2版本)[30]删去低质量读段,保留高质量的测序读段用于后续的分析.使用FastQC软件(0.11.9版本)[31]对下机数据进行质量控制.下载参考基因组和基因注释文件,使用HISAT2软件(2.0.5版本)[32]构建参考基因组的索引,并与参考基因组hg19进行比对;根据RefSeq转录本注释确定转录水平,并使用featureCounts程序(1.5.0-p3版本)[33]对能够比对到基因的读段数量进行统计,使用FPKM(fragments per kilobase of exon model per million mapped fragments)对表达量进行标准化,以使不同基因和样本之间的表达水平具有可比性;使用DESeq软件(1.39.0版本)[34]对不同组间的基因表达进行差异分析,差异表达基因(differentially expressed gene,DEG)被定义为至少一个重复的FPKM>1,且差异倍数(fold change,FC)对数的绝对值()>1,P<0.05.使用ClusterProfiler软件(4.10.1版本)对DEG进行功能富集分析[35].本研究中所有RNA-seq原始测序数据均已上传至国家生物信息中心(https://www.cncb.ac.cn/),GSA-Human编号为HRA009071. ...
1
... 所有的RNA-seq原始数据均以FASTQ格式下机.使用Cutadapt软件(1.9.1版本)去除接头序列[29],用Trimmomatic软件(0.3.2版本)[30]删去低质量读段,保留高质量的测序读段用于后续的分析.使用FastQC软件(0.11.9版本)[31]对下机数据进行质量控制.下载参考基因组和基因注释文件,使用HISAT2软件(2.0.5版本)[32]构建参考基因组的索引,并与参考基因组hg19进行比对;根据RefSeq转录本注释确定转录水平,并使用featureCounts程序(1.5.0-p3版本)[33]对能够比对到基因的读段数量进行统计,使用FPKM(fragments per kilobase of exon model per million mapped fragments)对表达量进行标准化,以使不同基因和样本之间的表达水平具有可比性;使用DESeq软件(1.39.0版本)[34]对不同组间的基因表达进行差异分析,差异表达基因(differentially expressed gene,DEG)被定义为至少一个重复的FPKM>1,且差异倍数(fold change,FC)对数的绝对值()>1,P<0.05.使用ClusterProfiler软件(4.10.1版本)对DEG进行功能富集分析[35].本研究中所有RNA-seq原始测序数据均已上传至国家生物信息中心(https://www.cncb.ac.cn/),GSA-Human编号为HRA009071. ...
1
... 所有的RNA-seq原始数据均以FASTQ格式下机.使用Cutadapt软件(1.9.1版本)去除接头序列[29],用Trimmomatic软件(0.3.2版本)[30]删去低质量读段,保留高质量的测序读段用于后续的分析.使用FastQC软件(0.11.9版本)[31]对下机数据进行质量控制.下载参考基因组和基因注释文件,使用HISAT2软件(2.0.5版本)[32]构建参考基因组的索引,并与参考基因组hg19进行比对;根据RefSeq转录本注释确定转录水平,并使用featureCounts程序(1.5.0-p3版本)[33]对能够比对到基因的读段数量进行统计,使用FPKM(fragments per kilobase of exon model per million mapped fragments)对表达量进行标准化,以使不同基因和样本之间的表达水平具有可比性;使用DESeq软件(1.39.0版本)[34]对不同组间的基因表达进行差异分析,差异表达基因(differentially expressed gene,DEG)被定义为至少一个重复的FPKM>1,且差异倍数(fold change,FC)对数的绝对值()>1,P<0.05.使用ClusterProfiler软件(4.10.1版本)对DEG进行功能富集分析[35].本研究中所有RNA-seq原始测序数据均已上传至国家生物信息中心(https://www.cncb.ac.cn/),GSA-Human编号为HRA009071. ...
1
... 所有的RNA-seq原始数据均以FASTQ格式下机.使用Cutadapt软件(1.9.1版本)去除接头序列[29],用Trimmomatic软件(0.3.2版本)[30]删去低质量读段,保留高质量的测序读段用于后续的分析.使用FastQC软件(0.11.9版本)[31]对下机数据进行质量控制.下载参考基因组和基因注释文件,使用HISAT2软件(2.0.5版本)[32]构建参考基因组的索引,并与参考基因组hg19进行比对;根据RefSeq转录本注释确定转录水平,并使用featureCounts程序(1.5.0-p3版本)[33]对能够比对到基因的读段数量进行统计,使用FPKM(fragments per kilobase of exon model per million mapped fragments)对表达量进行标准化,以使不同基因和样本之间的表达水平具有可比性;使用DESeq软件(1.39.0版本)[34]对不同组间的基因表达进行差异分析,差异表达基因(differentially expressed gene,DEG)被定义为至少一个重复的FPKM>1,且差异倍数(fold change,FC)对数的绝对值()>1,P<0.05.使用ClusterProfiler软件(4.10.1版本)对DEG进行功能富集分析[35].本研究中所有RNA-seq原始测序数据均已上传至国家生物信息中心(https://www.cncb.ac.cn/),GSA-Human编号为HRA009071. ...
1
... 首先使用Trimmomatic软件(0.3.2版本)[30]对原始测序数据进行质量控制,包括去除低质量的碱基、修剪接头序列和过滤掉低质量的读段;过滤掉的数据用FastQC软件(0.11.9版本)[31]进行质量控制;之后使用工具Bowtie2软件(2.5.2版本)将质控过的测序数据与参考基因组hg19进行比对[36],生成BAM文件,该文件包含了每个读段的比对位置信息.使用MACS2软件(2.1.4版本)识别基因组上染色质开放区域显著富集的位点或峰[37],并用MEME软件(5.5.7版本)[38]对这些峰信号进行注释;利用DiffBind软件(2.10版本)[39]对不同样本的ATAC-seq数据进行差异分析,以确定差异的染色质开放区域.利用基序(motif)的序列分析工具(MEME中的AME工具)进行相关富集分析,比对的基序数据库为JASPAR2022 CORE non-redundant v2,比对基因组为hg19.利用GREAT(Genomic Regions Enrichment of Annotations Tool)在线网站(http://great.stanford.edu/public/html/)对差异染色质开放区域进行功能富集分析,以了解这些区域所涉及的生物学过程、通路和功能.常用的富集分析工具包括GO(Gene Ontology)、KEGG(Kyoto Encyclopedia of Genes and Genomes)和GREAT等.本研究中所有ATAC-seq原始测序数据均已上传至国家生物信息中心(https://www.cncb.ac.cn/),GSA-Human编号为HRA009070. ...
1
... 首先使用Trimmomatic软件(0.3.2版本)[30]对原始测序数据进行质量控制,包括去除低质量的碱基、修剪接头序列和过滤掉低质量的读段;过滤掉的数据用FastQC软件(0.11.9版本)[31]进行质量控制;之后使用工具Bowtie2软件(2.5.2版本)将质控过的测序数据与参考基因组hg19进行比对[36],生成BAM文件,该文件包含了每个读段的比对位置信息.使用MACS2软件(2.1.4版本)识别基因组上染色质开放区域显著富集的位点或峰[37],并用MEME软件(5.5.7版本)[38]对这些峰信号进行注释;利用DiffBind软件(2.10版本)[39]对不同样本的ATAC-seq数据进行差异分析,以确定差异的染色质开放区域.利用基序(motif)的序列分析工具(MEME中的AME工具)进行相关富集分析,比对的基序数据库为JASPAR2022 CORE non-redundant v2,比对基因组为hg19.利用GREAT(Genomic Regions Enrichment of Annotations Tool)在线网站(http://great.stanford.edu/public/html/)对差异染色质开放区域进行功能富集分析,以了解这些区域所涉及的生物学过程、通路和功能.常用的富集分析工具包括GO(Gene Ontology)、KEGG(Kyoto Encyclopedia of Genes and Genomes)和GREAT等.本研究中所有ATAC-seq原始测序数据均已上传至国家生物信息中心(https://www.cncb.ac.cn/),GSA-Human编号为HRA009070. ...
1
... 首先使用Trimmomatic软件(0.3.2版本)[30]对原始测序数据进行质量控制,包括去除低质量的碱基、修剪接头序列和过滤掉低质量的读段;过滤掉的数据用FastQC软件(0.11.9版本)[31]进行质量控制;之后使用工具Bowtie2软件(2.5.2版本)将质控过的测序数据与参考基因组hg19进行比对[36],生成BAM文件,该文件包含了每个读段的比对位置信息.使用MACS2软件(2.1.4版本)识别基因组上染色质开放区域显著富集的位点或峰[37],并用MEME软件(5.5.7版本)[38]对这些峰信号进行注释;利用DiffBind软件(2.10版本)[39]对不同样本的ATAC-seq数据进行差异分析,以确定差异的染色质开放区域.利用基序(motif)的序列分析工具(MEME中的AME工具)进行相关富集分析,比对的基序数据库为JASPAR2022 CORE non-redundant v2,比对基因组为hg19.利用GREAT(Genomic Regions Enrichment of Annotations Tool)在线网站(http://great.stanford.edu/public/html/)对差异染色质开放区域进行功能富集分析,以了解这些区域所涉及的生物学过程、通路和功能.常用的富集分析工具包括GO(Gene Ontology)、KEGG(Kyoto Encyclopedia of Genes and Genomes)和GREAT等.本研究中所有ATAC-seq原始测序数据均已上传至国家生物信息中心(https://www.cncb.ac.cn/),GSA-Human编号为HRA009070. ...
1
... 首先使用Trimmomatic软件(0.3.2版本)[30]对原始测序数据进行质量控制,包括去除低质量的碱基、修剪接头序列和过滤掉低质量的读段;过滤掉的数据用FastQC软件(0.11.9版本)[31]进行质量控制;之后使用工具Bowtie2软件(2.5.2版本)将质控过的测序数据与参考基因组hg19进行比对[36],生成BAM文件,该文件包含了每个读段的比对位置信息.使用MACS2软件(2.1.4版本)识别基因组上染色质开放区域显著富集的位点或峰[37],并用MEME软件(5.5.7版本)[38]对这些峰信号进行注释;利用DiffBind软件(2.10版本)[39]对不同样本的ATAC-seq数据进行差异分析,以确定差异的染色质开放区域.利用基序(motif)的序列分析工具(MEME中的AME工具)进行相关富集分析,比对的基序数据库为JASPAR2022 CORE non-redundant v2,比对基因组为hg19.利用GREAT(Genomic Regions Enrichment of Annotations Tool)在线网站(http://great.stanford.edu/public/html/)对差异染色质开放区域进行功能富集分析,以了解这些区域所涉及的生物学过程、通路和功能.常用的富集分析工具包括GO(Gene Ontology)、KEGG(Kyoto Encyclopedia of Genes and Genomes)和GREAT等.本研究中所有ATAC-seq原始测序数据均已上传至国家生物信息中心(https://www.cncb.ac.cn/),GSA-Human编号为HRA009070. ...




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}